He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Invitro.umassmed.edu
Possible Role of Nitric Oxide and Arachidonic Acid Pathways in Hypoxia-Induced Contraction of Rabbit Coronary Artery Rings
C. Barbé1, V. De Crescenzo2, F. Diemont2 and P. Bonnet2
1Lab. de Neurophysiologie, Équipe du Préconditionnement du Myocarde, Université d’Angers, Cedex, France;2Lab. de Physiologie des Cellules Cardiaques et Vasculaires, CNRS UMR 6542, Faculté de Médecine, Tours, France
Abstract
In isolated coronary arteries, hypoxia induces an increase in
nary arteries of dogs (Borda et al., 1980; Vanhoutte, 1988),
tone by releasing an unidentified endothelium-derived con-
pigs (Rubanyi & Paul, 1985) and sheep (Kwan et al., 1989),
tracting factor (EDCF). Isometric force was measured in
it has been suggested that vasospasm could be triggered by
an isolated rabbit coronary artery ring at 37°C in control
a decrease in oxygen partial pressure. The vascular endothe-
and high K+ (40 mM) pre-contracted conditions. Hypoxia
lium modulates the tone of the underlying smooth muscle by
(15 mmHg pO2) induced by equilibrating the perfusate with
producing relaxing (Furchgott, 1983; Bolton & Clapp, 1986)
nitrogen. Hypoxia did not affect the resting tone but induced
and contracting factors (Vanhoutte, 1987). A modulation of
an endothelium-dependent contraction on pre-contracted
tone, secondary to the release of diffusible endothelium-
rings. Inhibitors of nitric oxide (NO) were tested, L-NAME
derived contracting factors (EDCF), can play a major role
(10-4 M) totally and L-NMMA (10-4 M) partially convert the
in coronary artery vasospasm and myocardial ischemia
hypoxic contraction to an hypoxic relaxation. The addition of
(Rubanyi & Vanhoutte, 1985; Pearson et al., 1991). EDCF
L-arginine (10-4 or 10-3 M) did not restore the response.
has not yet been identified (Rubanyi & Vanhoutte, 1985) nor
Methylene blue (10-5 M) and ODQ (1 H-[1,2,4] oxadiazolo-
quantified by bioassay. Previous studies have excluded cate-
[4,3-a] quinoxalin-1-one, 10-5 M), both inhibitors of guany-
cholamines, serotonin, histamine, adenine nucleotides and
late cyclase, also changed the hypoxic contraction into a
cyclooxygenase products as EDCF (De Mey & Vanhoutte,
hypoxic relaxation. Catalase (1200 U/ml), which decom-
1983; Rubanyi & Vanhoutte, 1985; Katusic & Vanhoutte,
poses hydrogen peroxide (H2O2), and superoxide dismutase
1986). The vasoconstrictive peptide, endothelin is an unlikely
(150 U/ml, SOD), a free radical scavenger, did not change the
candidate for EDCF because the hypoxic contraction is
hypoxic response but quinacrine (50 mM), an inhibitor of
observed quickly after the onset of the hypoxic exposure and
phospholipase A2, significantly decreased it. Inhibitors of
quickly reverses (Vanhoutte et al., 1989), whereas endothe-
arachidonic acid metabolism (indomethacin, diethylcarba-
lin needs time to be synthesised before being released. In
mazine, miconazole) however did not affect the hypoxic
human coronary arteries, indomethacin and diethylcarba-
response. We conclude that in K+ pre-contracted rabbit coro-
mazine suppress hypoxic contraction whereas indomethacin
nary artery rings, hypoxia induces a contraction which is
does not prevent the hypoxic contraction in dog coronary
nitric oxide and arachidonic acid dependent.
arteries. Three different mechanisms have been proposed toaccount for hypoxic contraction. There are the inhibition of
Keywords: hypoxic contraction, NO, arachidonic acid, coro-
an endothelium-derived relaxing factor (EDRF), for example
NO (Rees et al., 1989; Yang et al., 1994), the synthesis or therelease of contractile superoxide anions (Pearson et al., 1991;Lin et al., 1991) or the participation of an arachidonic acid
Introduction
(AA) metabolism product (Katusic & Vanhoutte, 1986; Rees
Vasospasm, one cause of myocardial infarction can be
et al., 1989; Graff & Gozal, 1999). In this study, we con-
defined as a sustained coronary artery contraction. As
firmed the role of the endothelium in the hypoxic contraction
hypoxia induces contraction in isolated rings of large coro-
and tested each of these three hypotheses by using nitric
Address correspondence to: Dr Bonnet, Lab. de Physiologie des Cellules Cardiaques et Vasculaires, CNRS UMR 6542, Faculté deMédecine, 2 bis bd Tonnellé, 37032 Tours, France. Tel.: +33 2 47 36 60 91; Fax: +33 2 47 36 60 64; E-mail: [email protected]
oxide synthase blockers, superoxide anion scavengers and
increasing doses of acetylcholine (ACh) in rings pre-
different blockers of AA metabolism. The goal of our study
contracted by histamine (His). This test was done at the end
was to focus on the pharmaco-physiological events explain-
of the 120-min equilibration period before the first hypoxic
ing the hypoxic contraction and the factors that determine
challenge and again at the end of the experiment. Relaxation
is known to occur only in preparations with intact endothe-lium (Furchgott & Zawadzki, 1980). This relaxation was usedas a positive test. Rings were sequentially superfused with
Material and methods
normoxic high K+ solution (40 mM) for 15 min, followed bysuperfusion with hypoxic high K+ solution until we obtained
Animal preparation
a maximal hypoxic contraction. We considered that the
Rabbits weighing 1.5 to 2 kg were heparinized (3500 UI) by
maximal hypoxic effect was reached when the initial increase
intraperitoneal injection and killed by cervical dislocation.
in tension remained stable for approximately five minutes.
After thoracotomy, the heart was quickly excised and placed
Rings were then superfused again with normoxic high K+
in a cold (4°C), calcium-free physiological saline solution
solution for 15 minutes. After this control hypoxic protocol,
with the following composition (in mmol/l): NaCl 138.6; KCl
the coronary artery rings were returned to baseline tension
5.4; MgCl2 1.2; NaH2PO4 0.33; HEPES 10; Glucose 11. The
by superfusion with normoxic PSS solution. After 60 min, the
pH was adjusted to 7.4 with NaOH. The epicardial part of the
same hypoxic protocol was repeated but in the presence of a
left circumflex coronary artery was removed, cleaned of
test drug dissolved in high K+ solution. The reproducibility
surrounding tissues taking care not to damage the luminal
of the hypoxic response was tested in control arterial rings
surface and prepared as segments (4–5 mm in length and 500–
by exposure to three successive hypoxic periods separated by
1000 mm in external diameter). In some rings the endothelium
20-min normoxia. We observed that repeated exposures to
was mechanically removed by rubbing the intima. Each
hypoxia neither potentiated nor inhibited of the responses
segment was suspended by two tungsten wires of 15 mm
of isolated coronary artery rings. A single drug was tested
of diameter passed through its lumen (internal diameter 400–
800 mm). One wire was anchored to the bottom of the organchamber and the other was connected to a force transducer
(Kent TRN 001–220) for measurement of isometric force. Segments of coronary artery, with or without endothelium,
Acetylcholine (ACh), Histamine (His), catalase, superoxide
from the same animal were suspended in four organ chambers
dismutase (SOD), quinacrine, diethylcarbamazine (DEC),
(6 ml volume) containing normoxic physiological saline solu-
miconazole, Nw-nitro-L-arginine methyl ester (L-NAME),
tion (PSS) which had the following composition (in mmol/l):
Nw-monomethyl L-arginine (L-NMMA), indomethacin,
NaCl 138.6; KCl 5.4; MgCl2 1.2; NaH2PO4 0.33; CaCl2 1.8;
methylene blue (MB) and 1H-[1,2,4]oxadiazolo[4,3-
HEPES 10; glucose 11. The pH was adjusted to 7.4 with
a]quinoxalin-1-one (ODQ) were obtained from SIGMA.
NaOH and the temperature held at 37°C. Before experiments,rings were preloaded (2 g) and allowed to equilibrate for 120
Statistical analysis
minutes at their optimal resting tension. Normoxic solutionswere equilibrated with 20% O2–80% N2 (pO2 150 mmHg).
The tension developed under hypoxia before and after the
Hypoxia was induced by switching to solution equilibrated
addition of each drug was compared. Contraction and relax-
with 100% N2. The pO2 of the hypoxic solution measured with
ation are expressed as % of the contraction induced by nor-
To test whether the ring preparation had an intact endothe-
All the results were expressed as mean ± standard error
lium, sections were taken from the coronary artery. Tissues
of the mean. The n value quoted is the number of rings used.
were fixed for a minimum of 48 h in 10% formalin and
Statistical evaluation of data in the same groups was per-
embedded in parafin wax. Sections of 4–5 mm were cut and
formed by paired Student’s t test analysis. Differences were
stained with haematoxylin and eosine. Using argentafin col-
considered significant when P was less than 0.05.
orimetry we demonstrated that the technical procedure main-tained an intact endothelial layer.
All animals received care in compliance with the ‘Guide
for the Care and Use of Laboratory Animals’ published by
Effects of hypoxia on ring tone
the National Institute of Health (NIH publication n°85-23,revised 1985).
Preliminary experiments determined the hypoxic response ofisolated rabbit coronary artery rings. In the first series ofexperiments, rings were superfused for 15 min with normoxic
Protocol
PSS followed by 15 min with hypoxic PSS. Hypoxia did not
The presence or the absence of a functional endothelium able
affect the resting tone (n = 11, Fig. 1A). The concentration
to release NO was tested by determining the response to
of K+ required to pre-constrict arterial rings, in order to
Isometric tension recording of an isolated rabbit coronary artery ring. A: Ring was exposed to hypoxic PSS solution. No effect
was observed. B: Ring was pre-contracted with 40 mM K+ solution (K+ 40 mM) before exposure to hypoxia. Hypoxic K+ solution induced anadditonal contraction of about 3 mN.
obtain an optimal reaction to hypoxia, was determined by
arginine (L-NMMA, 10-4 M), two inhibitors of NO synthase.
successive 15-min periods of superfusion of normoxic,
Hypoxia induced a contraction of 41.3 ± 12.3% (n = 8)
hypoxic and normoxic with different high K+ solutions
but in the presence of L-NAME, hypoxia induced a relax-
obtained by equimolar substitution of NaCl by KCl. In all
ation (43.5 ± 6.2% (n = 8)). The L-NAME mediated
cases (n = 6) hypoxia induced a further contraction of arte-
conversion of an hypoxic contraction into a relaxation was
rial rings pre-contracted by 40 mM K+ solution (Fig. 1B). In
not reversed by the addition of L-arginine at two concentra-
each case the effect of hypoxia was fully reversible and the
tions (10-4 and 10-3 M, n = 6 for each) (Fig. 3). The
level of contraction induced by high K+ solution post-hypoxia
amplitude of the contraction induced by high K+ solution was
was the same as that induced by high K+ pre-hypoxia (Fig.
significantly increased by L-NAME (4.9 ± 2.9 mN vs 23 ±
3.6 mN). This increase in high K+ contraction was partially
The effect of the endothelium was examined in 6 coro-
reversed by L-arginine since the magnitude of the K+ con-
nary artery rings where the endothelium had been removed
traction returned to 10.2 ± 3.1 mN. Experiments performed
mechanically and its absence verified by the failure of ACh
with L-NMMA (5 ¥ 10-5 and 10-4 M) modified the hypoxic
to induce relaxation (Fig. 2). The magnitude of the high K+
response: upon hypoxia a contraction was observed followed
contraction was the same (4.2 ± 2.1 mN vs 4.4 ± 1.9 mN in
by a relaxation (Fig. 4). The addition of L-arginine at 10-4
endothelium and endothelium-denuded rings, respectively)
and 10-3 M (n = 7) did not reverse the original hypoxic
suggesting that our technique to remove the intimal layer pre-
response (Fig. 4). The hypoxic contraction decreased from
served smooth muscle contraction capabilities. Hypoxia
4.2 ± 1.6 mN to 2.5 ± 0.9 mN (p < 0.05) in the presence of
induced a contraction of 34.8 ± 8.3% in rings with endothe-
L-NMMA. In the presence of L-NMMA with 10-3 M L-
lium but a relaxation of 37 ± 4.3% in rings without endo-
arginine the hypoxic contraction of 2.2 ± 0.9 mN was not
thelium (Fig. 2). In differents experiments rings were
different from the hypoxic contraction observed without L-
precontracted by histamine (10-5 M) (in place of high K+
arginine (2.5 ± 0.9 mN). L-NMMA increased the K+-
solution) with intact endothelium hypoxia still induced a con-
induced contraction (6.1 ± 2.8 mN control, 18.5 ± 6.2 mN
traction of 48 ± 7% and a relaxation of 26 ± 5% without
with L-NMMA) and this effect was partially reversed by the
addition of L-arginine (18.5 ± 6.2 mN to 7.8 ± 4.1 mN) sug-gesting that L-NMMA induced a blockade of the basal NO
Role of nitric oxide metabolism
Since NO is known to increase cGMP concentration in
The endothelium-dependent hypoxia-induced contraction
smooth muscle, we blocked its synthesis with methylene blue
has been suggested by Yang and co-workers (1994) to be the
(10-5 M), a potent inhibitor of guanylate cyclase. In the pres-
result of an inhibition in basal EDRF release. To test this
ence of methylene blue the magnitude of the high K+ con-
hypothesis, we blocked NO synthesis by Nw-nitro-L-arginine
traction did not change (6.8 ± 1 mN in control vs 9.6 ±
methyl ester (L-NAME, 10-4 M) and Nw-monomethyl L-
1.3 mN with the drug). Methylene blue changed the hypoxic
Isometric tension recording of isolated rabbit coronary artery rings with (A) or without (B) endothelium. Rings were pre-contracted
with either 10-5 M histamine (HIS) (left panel) or with 40 mM K+ solution (K+ 40 mM) (right panel). The presence of endothelium was testedon these rings by the response to increasing doses (a, b, c, d, respectively 10-7, 10-6, 10-5 and 10-4 M) of acetylcholine (left panel). Hypoxiainduced a contraction if the endothelium was intact (A) and a relaxation in a ring without endothelium (B).
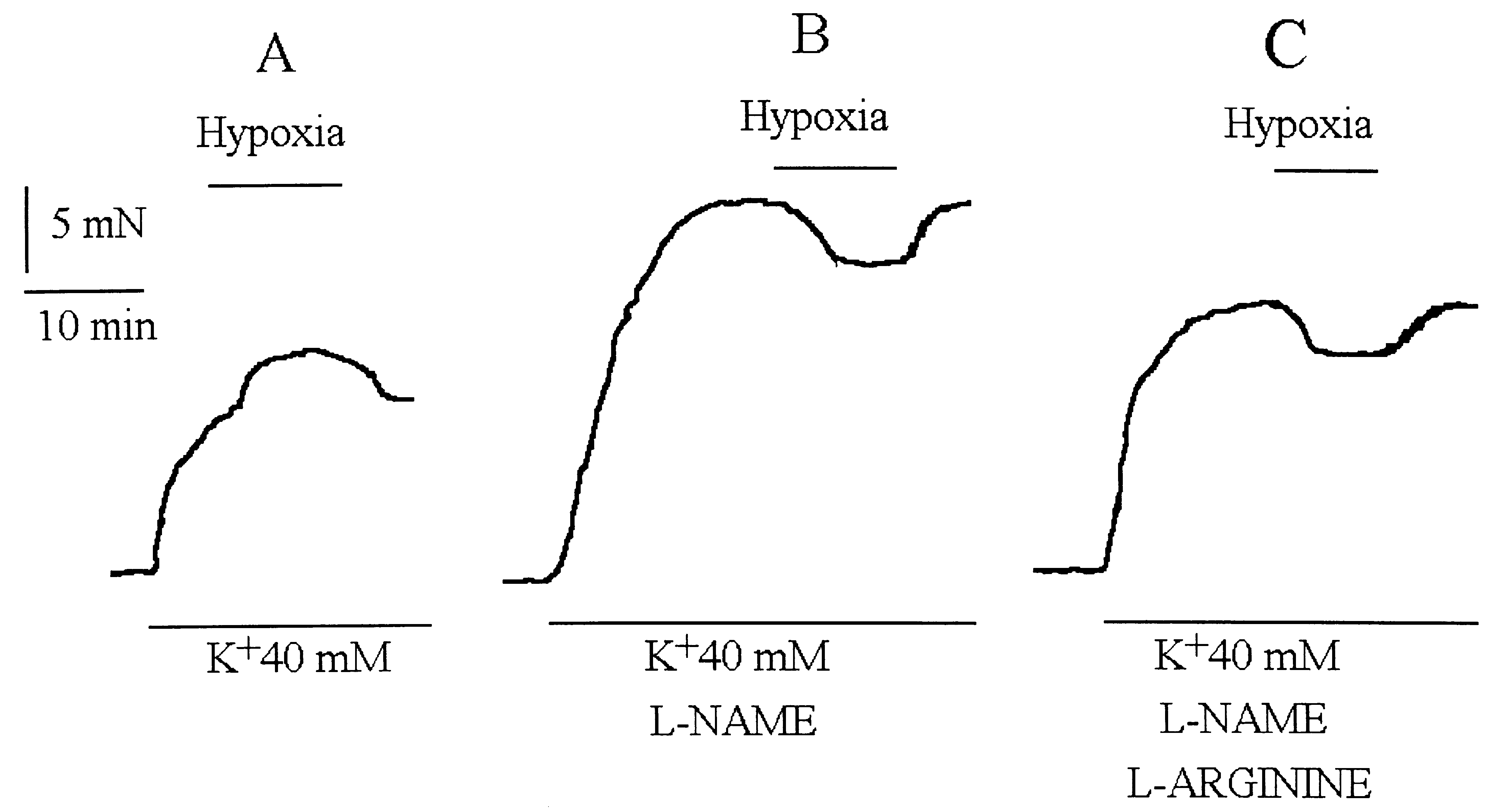
Isometric tension recording of an isolated rabbit coronary artery ring exposed to 40 mM hypoxic K+ solution. A: In control condi-
tions, hypoxia induced a contraction. B: In the presence of L-NAME (10-4 M) hypoxia induced a relaxation. C: The addition L-arginine (10-3 M) did not restore the hypoxic contraction.
contraction into a hypoxic relaxation (n = 7) (Table 1). After
tion into a hypoxic relaxation without any change in the high
washout, the hypoxic contraction was restored. We also
K+ contraction (Table 1). We conclude that hypoxic EDCF
tested ODQ (10-5 M) which is a more specific blocker of
release is dependent on the NO synthase pathway. Contrary
guanylate cyclase. ODQ also changed the hypoxic contrac-
to NO inhibitors, guanylate cyclase blockers did not enhance
Isometric tension recording of an isolated coronary artery ring pre-contracted by 40 mM K+ solution. A: In control conditions. B:
Exposure to L-NMMA (50 mM) (1) induced an additional contraction. The increase in L-NMMA concentration to 10-4 M (2) was withoutfurther effect on tension. Hypoxia induced a small hypoxic contraction followed by a relaxation. C: The application of L-NMMA (10-4 M)and L-arginine (10-3 M) (3) did not potentiate the potassium contraction as seen in B but hypoxia still induced the same response.
Effect of Methylene blue and ODQ on the hypoxic
Role of arachidonic acid metabolism
response express in percentage of the maximal amplitude of the
A role for arachidonic acid metabolism particularly
cyclooxygenase products has been proposed by severalauthors to explain the hypoxic contraction (Rubanyi &
Vanhoutte, 1985). The blockade of the endogenous synthesis
of arachidonic acid by 50 mM quinacrine an inhibitor of phos-pholipase A2 induced a significant decrease in the hypoxiccontraction (31 ± 6.5% vs 2.9 ± 1.7%, n = 6, p < 0.05)without any effect on high K+-induced contraction. After
wash-out, the hypoxic contraction reached the same value as
K+ contraction by 31 ± 15% and hypoxic contraction from9.5 ± 2.1 mN to 3.1 ± 1.9 mN, (p < 0.05). Since coronary
the K+ pre-contraction. According to the hypothesis of Yang
endothelial cells and coronary arteries metabolise arachi-
et al. (1994), we show that the NO pathway is implicated in
donic acid via the cyclooxygenase, lipooxygenase, and
cytochrome P-450 pathway, we blocked arachidonic acidmetabolism by using indomethacin (an inhibitor of cyclooxy-genase), diethylcarbamazine (an inhibitor of lipooxygenase)
Role of oxidative metabolism in hypoxic response
and miconazole (an inhibitor of cytochrome P-450). None of
Pearson et al. (1991) suggested that free radicals like the
these inhibitors modified the hypoxic contraction (Table 2).
superoxide anion (O-2) could react with NO and form
We conclude that hypoxic contraction is dependent upon
peroxynitrite (ONOO-). To test this hypothesis, we examined
arachidonic acid but not its metabolites.
the effects of superoxide dismutase (150 U/ml, SOD), known to scavenge O-2, on the hypoxic contraction. We also used catalase (1200 U/ml) which decomposes hydrogen
Discussion
peroxide (H2O2). Neither SOD nor catalase significantlymodified the hypoxic contraction: 19.7 ± 2% vs 18.5 ± 2%
This study confirmed that rabbit coronary artery rings need
(n = 9); 49 ± 9.4% vs 45 ± 11% (n = 8), respectively
to be pre-contracted in order to undergo contraction by
(p < 0.05). We conclude that oxidative metabolism is not
hypoxia, as reported previously (De Mey & Vanhoutte, 1983;
involved in the mechanical response of coronary artery ring
Rubanyi & Vanhoutte, 1985). We also showed that the
response to hypoxia was independent of whether pre-
Effect of hypoxia on rabbit coronary arteries in the pres-
never observed this difference in reversibility, even with
ence of blockers of arachidonic acid metabolism. The baseline con-
high dose of L-arginine. This adds to the evidence that
traction was obtained with 40 mM potassium.
hypoxia might act on the binding sites between NOS and L-arginine. This effect seems to occur only during hypoxia
and not during normoxia as suggested by the reversibility of NOS inhibitors on the K+ contraction before hypoxic
We demonstrated that MB and ODQ converted hypoxia-
induced endothelium-dependent contraction to relaxation
without impairment of K+ induced contraction. It is well
Data are expressed as means ± SEM. DEC: diethylcarbazemine.
known that MB, like ODQ inhibits soluble guanylate cyclaseand thus depletes cGMP in vascular smooth muscle. Deple-tion of cGMP should lead to contraction (Murad et al., 1978;
contraction was evoked by superfusion with high K+ solution
Ignarro & Kadowitz, 1985). Gräser and Vanhoutte (1991)
demonstrated that cGMP was involved in the endothelium-
In agreement with previous studies we demonstrated that
dependent increase in tension caused by hypoxia and con-
hypoxic contraction of pre-contracted arteries depended on
cluded that a moderate increase in the cGMP level in the
the presence of the endothelium (Borda et al., 1980;
vascular smooth muscle cell is necessary for the hypoxic con-
Furchgott & Zawadski, 1980; De Mey & Vanhoutte, 1983;
traction in canine coronary artery. We suggest that hypoxia
Rubanyi & Vanhoutte, 1985; Kwan et al., 1989). This
acts on NOS to produce NO in a very specific concentration
hypoxic contraction might be due to a ‘down-regulation’ of
range that induces a moderate cGMP increase.
basal EDRF release which would lead to contraction (Rees
Pearson et al. (1991) tested the idea that NO could
et al., 1989). This hypothesis is supported by studies demon-
combine with other compounds, and found that superoxide
strating that hypoxia impairs the release of EDRF in canine
anion (O-2) formed in hypoxic endothelial cells combined
femoral artery (De Mey & Vanhoutte, 1983) and in canine
with NO to form the peroxynitrite anion (ONOO-) which was
coronary artery (Muramatsu et al., 1992). Our results show
then metabolized to an endothelium-derived contracting
an increase in the high K+ contraction secondary to L-NAME
factor or induced the synthesis of such a factor. However,
or L-NMMA exposure. This effect was not totally reversed
peroxynitrite anion itself could not reproduce the hypoxic
by L-arginine when using L-NAME but totally reversed with
contraction, and several authors (Liu et al., 1994; Wu et al.,
L-NMMA. This increase in high K+ contraction, secondary
1994) found that this compound had a vasodilator activity.
to NOS block exposure confirms a basal release of NO and
Peroxynitrite anion is a powerful oxidant which can react
strongly suggests that hypoxic contraction cannot be sec-
with a wide variety of compounds leading to the formation
ondary to the inhibition of basal EDRF release. Similar
of the endothelium derived-contracting factor. Our results
results were observed by other authors with different tissues
clearly demonstrated that free radicals were not involved in
and species (Pearson et al., 1991; Muramatsu et al., 1992;
the hypoxic contraction of rabbit coronary arteries.
Auch-Schwelk et al., 1992). This implicates the secretion
Cytochrome P-450 metabolites could play a role in the
of a contracting factor by hypoxia. We found a differential
hypoxic response (Campbell et al., 1996). However, since we
effect between L-NAME and L-NMMA. Our results clearly
did not observe any effect of miconazole, it seems unlikely
demonstrate that L-NAME reverses the hypoxic effect: a
that structural modification of cytochrome P-450 metabolites
hypoxic relaxation instead of a hypoxic contraction. In the
induced by hypoxia might explain the hypoxic contraction.
presence of L-NMMA we observed a transient hypoxic con-
Zelenkov et al. (1993) showed that indomethacin made no
traction followed by a relaxation. These two effects clearly
significant difference in hypoxic contraction of rat aorta,
indicate the obligatory role of NO in hypoxic contraction.
ruling out a prostaglandin mediator. The lack of effect of
As we found a differential effect with NOS inhibitors, we
inhibitors of the arachidonic acid metabolism pathway in our
propose that the blocking effect of L-NAME could be more
study leads to the same conclusion for rabbit coronary arter-
efficient under hypoxia compared to the blocking effect of L-
ies. On the contrary, quinacrine, an inhibitor of phospholi-
NMMA. This hypothesis is supported by a study that clearly
pase A2, inhibited the hypoxic contraction. Quinacrine is
demonstrated that relaxation induced by NO on vessels is
suspected to inhibit calcium channels (Nagano et al., 1996)
dose-dependent (Lincoln et al., 1996). Since both inhibitors
which could explain the decrease in the K+ pre-contraction
have the same blocking effect on the basal NO release under
with 10-3 M quinacrine. However, this phenomenon was not
normoxic conditions (Knowles & Moncada, 1994), we would
observed with 50 mM quinacrine which still inhibited the
suggest that hypoxia acts on the link between NOS and L-
hypoxic contraction. This strongly suggests that arachidonic
arginine or its competitive inhibitor (L-NAME and L-
acid itself and not its metabolites might be involved in the
NMMA). L-NAME, unlike L-NMMA, shows a progressive
and irreversible or slowly reversible inhibition of NOS
The NO and eicosanoid-producing metabolic pathways
(Knowles & Moncada, 1994). Under hypoxic conditions we
have been studied extensively. NO is an oxidizing radical that
has been reported to induce transcription factors (Herdegen
Gräser T, Vanhoutte PM (1991): Hypoxic facilitation of canine
et al., 1994) and interact with heme-containing proteins of
coronary arteries: role of the endothelium and cGMP. Am
guanylate cyclase (Lei et al., 1992). Guanylate cyclase acti-
J Physiol 261: H1769–H1777.
vation leads to an increase in cGMP-stimulated protein
Herdegen T, Rüdiger S, Mayer B, Bravo R, Zimmermann M
kinase G. Arachidonic acid is a contracting agent, which
(1994): Expression of nitric oxide synthase and colocalisa-
inhibits myosin light chain phosphatase (Parsons et al.,
tion with Jun, Fos and Krox transcription factors in spinal
1996) but it can also be a cofactor of protein kinase C like
cord neurons following noxious stimulation of the rat hind-
pare. Molec Brain Res 22: 245–258.
Our results implicate NO and eicosanoid-producing meta-
Ignarro LJ, Kadowitz PJ (1985): The pharmacological and phy-
bolic pathways in the hypoxic contraction of rabbit coronary
siological role of cyclic GMP in vascular smooth muscle
artery rings. We suggest that hypoxia induces a release of NO
relaxation. Ann Rev Pharmacol Toxicol 25: 171–191.
leading to a sufficient cGMP level necessary to be implicated
Katusic ZS, Vanhoutte PM (1986): Anoxic contractions in iso-
in the production of the EDCF, issued from the phospholi-
lated canine cerebral arteries: contribution of endothelium-
derived factors, metabolites of arachidonic acid, and
We conclude that hypoxic contraction observed in isolated
calcium entry. J Cardiovasc Pharmacol 8: S97–S101.
rabbit coronary artery rings depends upon arachidonic acid
Knowles RG, Moncada S (1994): Nitric oxide synthases in
and nitric oxide, and involves a cGMP-dependent intracellu-
mammals. Biochem J 298: 249–258.
lar mechanism. This mechanism may be implicated in coro-
Kwan YW, Wadsworth RM, Kane KA (1989): Effects of hypoxia
on the pharmacological responsiveness of isolated coronaryrings from the sheep. Br J Pharmacol 96: 849–856.
Lei SZ, Pan ZH, Aggarwal SK, Chen HSV, Hartman J, Sucher
NJ, Lipton SA (1992): Effect of nitric oxide production on
Acknowledgements
the redox modulatory site of the NMDA receptor-channel
This study supported by operating grants from the Fondation
complex. Neuron 8: 1087–1099.
de France. The authors thank May Fuentes for her help
Lin PJ, Pearson PJ, Cartier R, Schaff HV (1991): Superoxide
to type the manuscript and Manuel Rebocho for his techni-
anion mediates the endothelium-dependent contractions to
cal assistance. We thank Dr Kumar for checking the
serotonine by regenerated endothelium. J Thorac Cardio-
Lincoln TM, Cornwell TL, Komalavilas P, Macmillan-Crow LA,
Boerth A (1996): The nitric oxide-cyclic GMP signaling
References
system of biochemistry of smooth muscle contractionextract. In: Barany M, ed., Biochemistry of Smooth Muscle
Auch-Schwelk W, Katusic ZS, Vanhoutte PM (1992): Nitric
Contraction Extract. London, Academic Press, pp.
oxide inactivates endothelium derived- relaxing factor in
the rat aorta. Hypertension 19: 442–445.
Liu S, Beckman JS, Ku DD (1994): Peroxynitrite a product of
Bolton TB, Clapp LH (1986): Endothelial-dependent relaxant
superoxide and nitric oxide, produces coronary
actions of carbachol and substance P in arterial smooth
vasorelaxation in dogs. J Pharmacol Exp Ther 268: 1114–
muscle. Br J Pharmacol 87: 713–723.
Borda LJ, Shuchleib R, Henry PD (1980): Hypoxic contraction
Murad FC, Mittad CK, Arnold WP, Katsuki S, Kimura H (1978):
of isolated canine coronary artery. Mediation by potassium-
Guanylate cyclase: activation by azide, nitro compounds,
dependent exocytosis of norepinephrine. Circ Res 46:
nitric oxide, and hydroxyl radical and inhibition by hemo-
globin and myoglobin. Adr Cyclic Nucl Res 9: 145–158.
Campbell WB, Gebremedhin D, Pratt PF, Harder DR (1996):
Muramatsu M, Iwama Y, Shimizu K, Asano H, Toki Y, Miyasaki
Identification of epoxyeicosatrienoic acids as endothelium-
Y, Okumura K, Hashimoto H, Ito T (1992): Hypoxia-
derived hyperpolarizing factors. Circ Res 78: 415–423.
elicited contraction of aorta and coronary artery via
De Mey JG, Vanhoutte PM (1983): Anoxia and endothelium-
removal of endothelium-derived nitric oxide. Am J Physiol
dependent reactivity of the canine femoral artery. J Physiol
Nagano N, Imaizumi Y, Watanabe M (1996): Novel blockade of
Furchgott RF (1983): Role of endothelium in responses of vas-
Ca2+ current by quinacrine in smooth muscle cells of the g-
cular smooth muscle. Circ Res 53: 557–573.
uinea pig. Jpn J Pharmacol 71: 51–60.
Furchgott RF, Zawadzki JV (1980): The obligatory role of
Parsons SJW, Sumner MJ, Garland CJ (1996): Phospholipase A2
endothelial cells in the relaxation of arterial smooth muscle
and protein kinase C contribute to myofilament sensitiza-
by acetylcholine. Nature 299: 373–376.
tion to 5-HT in the rabbit mesenteric artery. J Physiol 491:
Graff GR, Gozal D (1999): Cardiorespiratory responses
to interleukin-1b in adult rats: role of nitric oxide, eicona-
Pearson PJ, Lin PJ, Schaff HV (1991): Production of
soids, and glucocorticoids. Arch Physiol Biochem 107: 97–
endothelium-derived contracting factor is enhanced after
coronary reperfusion. Ann Thorac Surg 51: 788–793.
Rees DD, Palmer RMJ, Moncada S (1989): Role of
endothelium-derived nitric oxide in the regulation of bloodpressure. Proc Nat Acad Sci USA 86: 3375–3378.
Rubanyi GM, Paul RJ (1985): Two distinct effects of oxygen on
vascular tone in isolated porcine coronary arteries. Circ Res56: 1–10.
Rubanyi GM, Vanhoutte PM (1985): Hypoxia releases a vaso-
constriction substance from the canine vascular endothe-lium. J Physiol 364: 45–56.
Vanhoutte PM (1987): Endothelium-dependent contractions in
arteries and veins. Blood Vessels 24: 141–144.
Vanhoutte PM (1988): The endothelium modulator of vascular
smooth muscle tone. N Engl J Med 319: 512–513.
Vanhoutte PM, Auch-Schwelk W, Boulanger C, Janssen PA,
Katusic ZS, Komori K, Miller VM, Schini VB Vidal M(1989): Does endothelin-1 mediate endothelium-dependentcontractions during anoxia? J Cardiovasc Pharmacol 13:S124–S128.
Wu M, Pritchard KAJR, Kamiznski PM, Fayngersh RP, Hintze
TH, Wolin MS (1994): Involvement of nitric oxide andnitrosothiols in relaxation of pulmonary arteries to peroxy-nitrite. Am J Physiol 266: H2108-H2113.
Yang XP, Wu ZJ, Xu YF, Dong W, Yang W, Fu SX, Li YS (1994):
Effects of inhibitor of endothelium-derived relaxing factoron hypoxic contraction of isolated pig coronary artery. ActaPharmacol Sin 15: 323–326.
Zelenkov P, McLoughlin T, Johns RA (1993): Endotoxin
enhances hypoxic contraction of rat aorta and pulmonaryartery through induction of EDRF/NO synthase. Am JPhysiol 265: L346-L354.
American Journal of Botany 87(5): 748–751. 2000. DISCOVERY OF DISTYLY IN N ARCISSUS (AMARYLLIDACEAE) 1 JUAN ARROYO2 AND SPENCER C. H. BARRETT3,42Departamento de Biologı´a Vegetal y Ecologı´a, Universidad de Sevilla, Apartado 1095, E-41080 Sevilla, Spain; and3Department of Botany, University of Toronto, 25 Willcocks Street, Toronto, Ontario, Canada, M5S 3B2There has been disagreem
Richard J. Fruncillo, M.D., Ph.D. 7 Lakeview Place Newtown Square, PA 19073 CURRICULUM VITAE EDUCATION Mercy Catholic Medical Center, Darby, PA Thomas Jefferson University, Philadelphia, PA The Hahnemann Medical College, Philadelphia, PA The Hahnemann Medical College Hospital, Philadelphia, PA The Hahnemann Medical College, Philadelphia, PA The University of Pennsylvania,
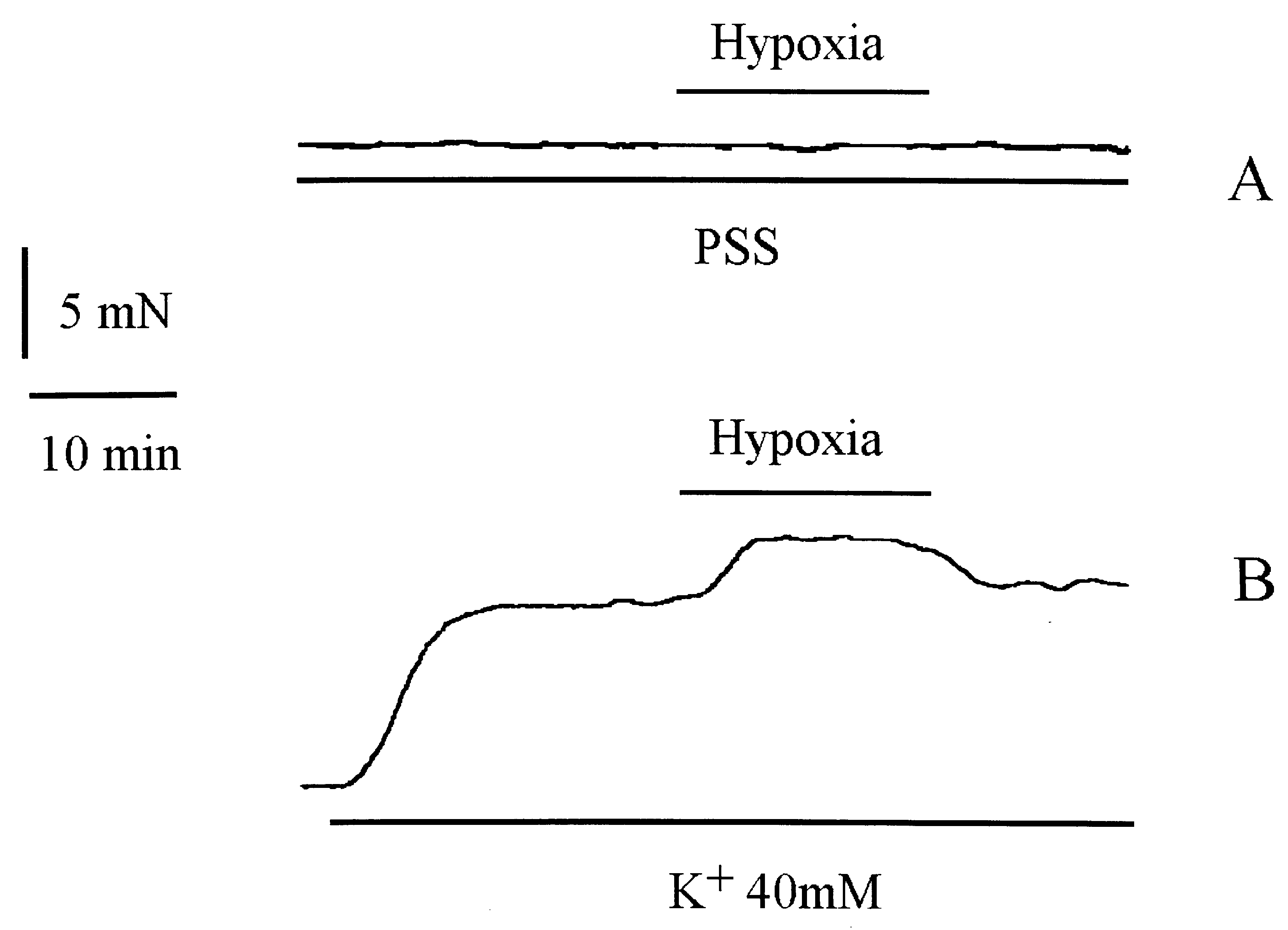 Isometric tension recording of an isolated rabbit coronary artery ring. A: Ring was exposed to hypoxic PSS solution. No effect
was observed. B: Ring was pre-contracted with 40 mM K+ solution (K+ 40 mM) before exposure to hypoxia. Hypoxic K+ solution induced anadditonal contraction of about 3 mN.
Isometric tension recording of an isolated rabbit coronary artery ring. A: Ring was exposed to hypoxic PSS solution. No effect
was observed. B: Ring was pre-contracted with 40 mM K+ solution (K+ 40 mM) before exposure to hypoxia. Hypoxic K+ solution induced anadditonal contraction of about 3 mN.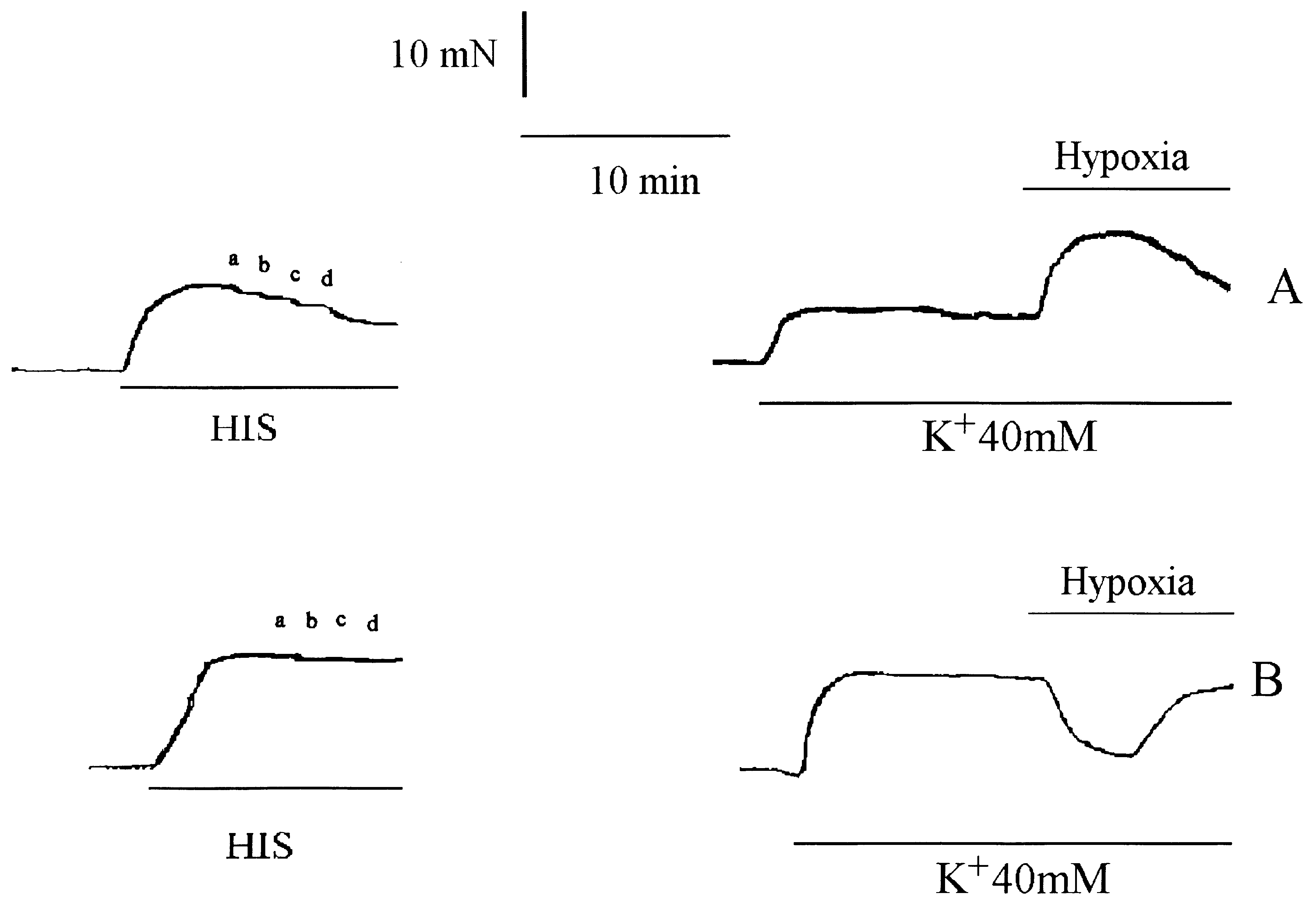
 Isometric tension recording of isolated rabbit coronary artery rings with (A) or without (B) endothelium. Rings were pre-contracted
with either 10-5 M histamine (HIS) (left panel) or with 40 mM K+ solution (K+ 40 mM) (right panel). The presence of endothelium was testedon these rings by the response to increasing doses (a, b, c, d, respectively 10-7, 10-6, 10-5 and 10-4 M) of acetylcholine (left panel). Hypoxiainduced a contraction if the endothelium was intact (A) and a relaxation in a ring without endothelium (B).
Isometric tension recording of isolated rabbit coronary artery rings with (A) or without (B) endothelium. Rings were pre-contracted
with either 10-5 M histamine (HIS) (left panel) or with 40 mM K+ solution (K+ 40 mM) (right panel). The presence of endothelium was testedon these rings by the response to increasing doses (a, b, c, d, respectively 10-7, 10-6, 10-5 and 10-4 M) of acetylcholine (left panel). Hypoxiainduced a contraction if the endothelium was intact (A) and a relaxation in a ring without endothelium (B).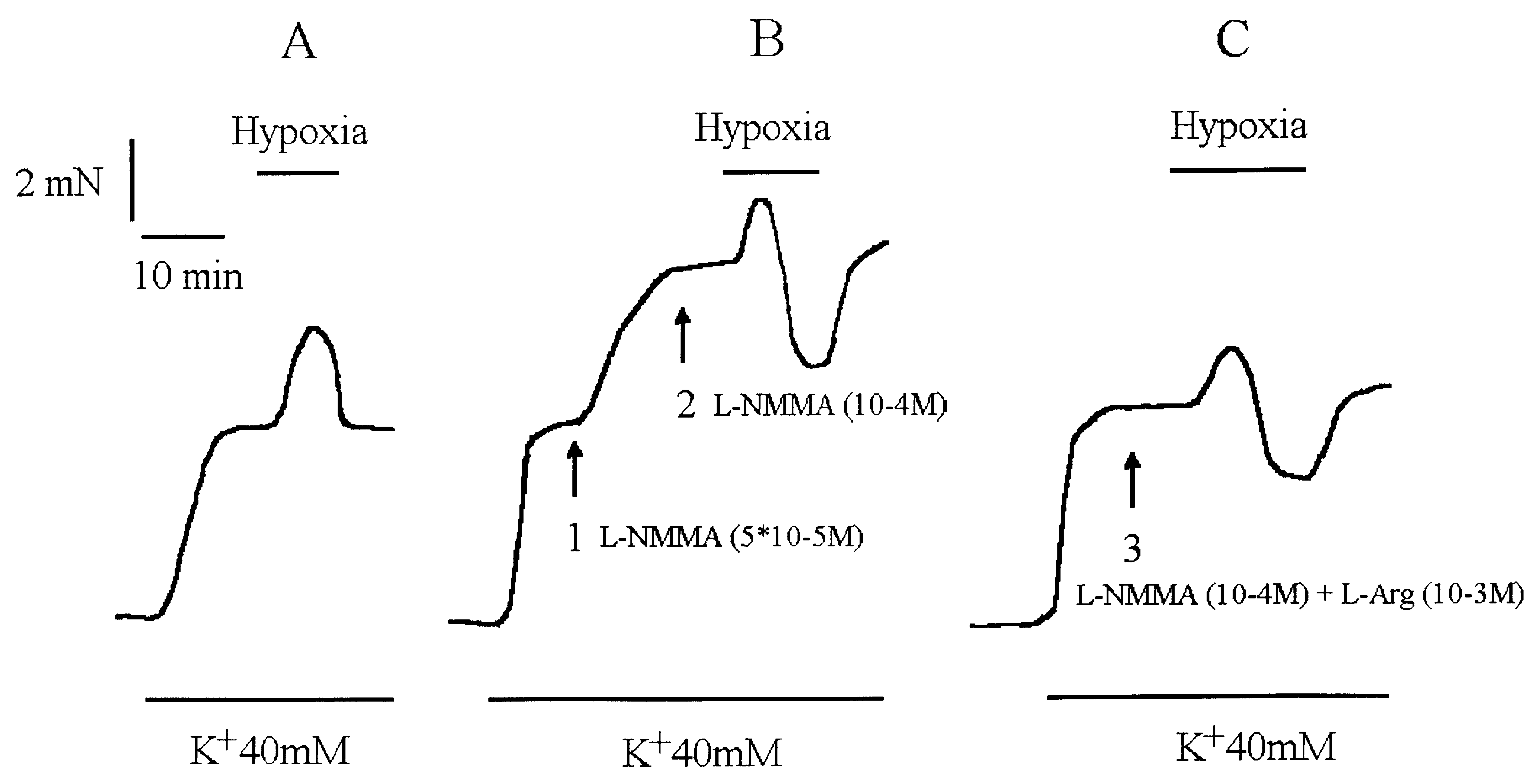 Isometric tension recording of an isolated coronary artery ring pre-contracted by 40 mM K+ solution. A: In control conditions. B:
Exposure to L-NMMA (50 mM) (1) induced an additional contraction. The increase in L-NMMA concentration to 10-4 M (2) was withoutfurther effect on tension. Hypoxia induced a small hypoxic contraction followed by a relaxation. C: The application of L-NMMA (10-4 M)and L-arginine (10-3 M) (3) did not potentiate the potassium contraction as seen in B but hypoxia still induced the same response.
Isometric tension recording of an isolated coronary artery ring pre-contracted by 40 mM K+ solution. A: In control conditions. B:
Exposure to L-NMMA (50 mM) (1) induced an additional contraction. The increase in L-NMMA concentration to 10-4 M (2) was withoutfurther effect on tension. Hypoxia induced a small hypoxic contraction followed by a relaxation. C: The application of L-NMMA (10-4 M)and L-arginine (10-3 M) (3) did not potentiate the potassium contraction as seen in B but hypoxia still induced the same response.