He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Abot_87_516.748_75
American Journal of Botany 87(5): 748–751. 2000. DISCOVERY OF DISTYLY IN NARCISSUS (AMARYLLIDACEAE)1
JUAN ARROYO2 AND SPENCER C. H. BARRETT3,4
2Departamento de Biologı´a Vegetal y Ecologı´a, Universidad de Sevilla, Apartado 1095, E-41080 Sevilla, Spain; and
3Department of Botany, University of Toronto, 25 Willcocks Street, Toronto, Ontario, Canada, M5S 3B2
There has been disagreement concerning the nature of sexual polymorphisms in Narcissus, a genus of insect-pollinated
geophytes native to the Mediterranean. The existence of trimorphic heterostyly in the genus has recently been confirmed,but the occurrence of distyly remains enigmatic. All sexually dimorphic species previously investigated possess two distinctstyle lengths but anthers of similar height. Stigma-height dimorphism does not qualify as true distyly because of the absenceof a reciprocal correspondence in stigma and anther position in the floral morphs. Such reciprocal herkogamy is generallyregarded as the defining feature of heterostyly. Here we report on distyly in N. albimarginatus (section Apodanthae), a rarespecies confined to a single mountain in northwestern Morocco. A population composed of equal numbers of long- andshort-styled plants exhibited reciprocal herkogamy with lower anthers of the long-styled morph and upper anthers of theshort-styled morph corresponding in height to stigmas of short- and long-styled plants, respectively. The presence of bothstigma-height dimorphism and distyly in Narcissus is of general significance to theoretical models of the evolution ofheterostyly. Key words:
Amaryllidaceae; distyly; Lloyd and Webb model; Narcissus; reciprocal herkogamy; stigma-height dimor-
Heterostyly has originated independently in at least 28
lesworth (1979) also investigated the stability of stigma-
animal-pollinated flowering plant families but the evo-
height dimorphism and found that it was difficult to
lutionary build-up of the polymorphism is still poorly
maintain in populations. Whether stigma-height dimor-
understood (Ganders, 1979a; Barrett, 1992). Theoretical
phism is an intermediate stage in the evolution of distyly
models differ in both the selective mechanisms invoked
has remained difficult to assess because of the absence
and in the sequence in which the morphological and
of comparative data. One source of evidence in favor of
physiological components of the syndrome are assembled
the Lloyd and Webb (1992a, b) model would be the co-
(Charlesworth and Charlesworth, 1979; Lloyd and Webb,
occurrence of stigma-height dimorphism and distyly
1992a, b; Richards, 1998). Unfortunately, little compar-
ative information is available to evaluate the different
Narcissus (Amaryllidaceae) is composed of ϳ40 spe-
pathways by which heterostyly has evolved and there
cies of insect-pollinated geophytes native to the Mediter-
have been few experimental studies that have examined
ranean. Populations are either monomorphic, dimorphic,
the contrasting predictions of theoretical models (Kohn
or trimorphic for style length (Barrett, Lloyd, and Arroyo,
and Barrett, 1992; Stone and Thomson, 1994).
1996) with at least a dozen species in three sections of
Lloyd and Webb (1992a, b) argued that the first stage
the genus (Apodanthae, Jonquillae, and Tazettae) pos-
in the evolution of distyly from an approach herkoga-
sessing length dimorphism of styles (Dulberger, 1964;
mous ancestor involves the establishment of a polymor-
Arroyo and Dafni, 1995; Baker, Thompson, and Barrett,
phism for stigma height but not anther height. They sug-
2000a, b). All species with stylar dimorphism possess
gested that the rarity of stigma-height dimorphism in het-
two anther levels within a flower, but these are positioned
erostylous groups is because this stage was rapidly passed
similarly at the upper portion of the floral tube in both
through during selection for complete reciprocal herko-
morphs [for example, see Fig. 1 in Dulberger (1964) and
gamy. Selection for differentiation in anther height would
Baker, Thompson, and Barrett, (2000a)]. Because of the
likely increase the proficiency of pollen dispersal between
absence of reciprocal herkogamy, dimorphic species of
morphs and be strongly selected in populations with only
Narcissus should not be considered distylous but instead
a dimorphism in stigma height. Charlesworth and Char-
are best described as possessing stigma-height dimor-phism (reviewed in Barrett, Jesson, and Baker, 2000). In
1 Manuscript received 19 August 1999; revision accepted 20 January
N. triandrus (section Ganymedes) three distinct style
The authors thank David Lloyd and Lawrence Harder for valuable
morphs commonly occur within populations and accom-
´ n, and J. A. Mejı´as for help in the field;
panying the differences in stigma height are three discrete
M. Ater (University of Tetouan) for logistic support; Bill Cole for prep-
anther levels. Because of the reciprocal nature of stigma
aration of the figures; Angela Baker for permission to cite unpublished
and anther heights, Barrett et al. (1997) described this
data; and research grants from DGESIC Spain (PB95-551, PB98-1144)
species as tristylous. In contrast to most heterostylous
4 Author for correspondence (FAX 416-978-5878, e-mail: barrett@
groups, heterostyly is rare in Narcissus whereas stigma-
height dimorphism is common, implying that there are
TABLE 1. Mean and (standard deviation) of stigma and anther heights
(mm) in the two floral morphs of distylous Narcissus albimargin-atus, a rare species from northwestern Morocco. See text for de-tails.
the nature of floral variation in the population a single flower was sam-pled from all individuals that were in anthesis and flowers were pre-served in 70% ethanol. The sample involved 97 plants, of which 88produced solitary-flowered inflorescences, eight had two flowers, andone had three flowers per inflorescence. Narcissus albimarginatus pos-
Distyly in Narcissus albimarginatus. Range of variation in
sesses pale-yellow pendulous flowers with a floral tube and a prominent
stigma (r) and anther heights (upper-level stamens ᭡, lower-level sta-
corona with a white margin. No pollinators were observed visiting the
mens ⅜) in a sample of 97 flowers from a population at Jebel Bou-
hachem mountain in northwestern Morocco. Note the reciprocal corre-spondence in height of stigmas of the long- and short-styled morphs
On return to the laboratory the style length (including stigma) and
with the upper- and lower-level stamens of the short- and long-styled
height of upper- and lower-level stamens (to anther base) of all flowers
morphs, respectively. Flowers are ranked by style length.
were measured using digital calipers to the nearest 0.1 mm. All mea-surements were made from the top of the ovary, and all stamens weremeasured and the average value taken for each stamen level. Data from
strong constraints on the evolution of reciprocal herko-
N. albimarginatus were then compared with published measurements
gamy in populations with stigma-height dimorphism. In
of sex-organ variation in seven Narcissus species with stigma-height
addition, there is no evidence for the occurrence of dial-
dimorphism to assess variation in the degree of reciprocity in stigmasand anthers.
lelic incompatibility or ancillary polymorphisms of pol-len and stigmas in any Narcissus species (Dulberger,1964; Arroyo and Dafni, 1995; Barrett, Lloyd, and Ar-
royo, 1996; Sage et al., 1999; Baker, Thompson, and Bar-
The population of N. albimarginatus was composed of
two discrete floral morphs differing in style length (Fig.
Here we provide quantitative evidence for the exis-
1). The population contained 45 long-styled plants and
tence of distyly in a rare species of Narcissus from north-
52 short-styled plants. This ratio does not differ signifi-
western Morocco. Narcissus albimarginatus D. & U.
cantly from equality (2 ϭ 0.505; P ϭ 0.477). Of partic-
Muller-Doblies was first described in 1989 from near the
ular interest was the finding that the position of the two
summit of the mountain Jebel (as in the original descrip-
stamen levels within a flower differed significantly be-
tion) Bouhachem (Muller-Doblies and Muller-Doblies,
tween the morphs, demonstrating that the population does
1989). The illustrations of N. albimarginatus accompa-
not exhibit a simple stigma-height dimorphism (Table 1).
nying the taxonomic description indicate two floral
In the long-styled morph the two stamen levels were in-
morphs differing in style length and anther position, and
cluded within the corona (upper-level stamens) and floral
the authors describe the species as ‘‘heterostylic.’’ How-
tube (lower-level stamens), whereas in the short-styled
ever, population samples were not reported and many au-
morph both stamen levels were exserted beyond the co-
thors have described Narcissus species as heterostylous
rona. Reciprocal herkogamy occurs in N. albimarginatus
when in fact the species exhibit stigma-height dimor-
because lower-level stamens in the long-styled morph and
phism not distyly (e.g., Henriques, 1887; Fernandes,
upper-level stamens of the short-styled morph correspond
1964; Webb, 1980; Valde´s, 1987; Richards, 1997; and see
in height to stigmas of the short- and long-styled morphs,
Barrett, Lloyd, and Arroyo, 1996, for a review). Because
respectively (Table 1). Because of this reciprocal corre-
of this problem we decided to investigate sex-organ var-
spondence in stigma and anther height N. albimarginatus
iation in N. albimarginatus further by attempting to lo-
cate the population described by D. and U. Muller-Dob-
The degree of stigma-anther reciprocity in N. albimar-
lies in Morocco and conducting detailed sampling to de-
ginatus is much greater than in Narcissus species with
termine whether the species was indeed truly distylous.
stigma-height dimorphism (Fig. 2). This is largely be-cause lower-level stamens in the long-styled morph cor-
respond more closely in height to stigmas of the short-
On 18 April 1999 the Djebel Bouhachem population was located in
styled morph, whereas in species with stigma-height di-
the Western Rif mountain range of northwestern Morocco. The popu-
morphism these stamens are usually positioned higher in
lation occurs along a sandstone ridge (not limestone as earlier reported)
the floral tube. Interestingly, the mean difference in stig-
at 1500 m growing under Cedrus atlantica forest and was composed of
ma height between the floral morphs in N. albimarginatus
ϳ100 flowering individuals in an approximate area of 200 ϫ 100 m.
is the largest of the eight dimorphic species of Narcissus
At the time of the visit the population was past peak flowering, but a
illustrated in Fig. 2. Disruptive selection on stigma height
sufficient number of flowers was available for sampling. To determine
during the evolution of reciprocal herkogamy may have
mechanisms that might favor the reverse evolutionary se-quence since this would likely reduce the proficiency ofcross-pollen dispersal between the floral morphs. Second,the widespread occurrence of stigma-height dimorphismin Narcissus, including species with varying degrees ofreciprocal herkogamy (Fig. 2), implies that variation ex-ists for the selection of distyly under the appropriate pol-lination environment. Finally, the isolated occurrence ofdistyly in a species apparently restricted to a single moun-tain in northwestern Morocco seems an unlikely place tolocate the ancestral polymorphic condition in Narcissus,unless of course this taxon represents a relictual speciesand all other distylous taxa have gone extinct. Clearly, aphylogeny of the genus would be required to fully resolvethese issues. In the meantime it seems to us that theweight of evidence favors the hypothesis that distyly inNarcissus is derived from stigma-height dimorphismthrough selection for more proficient cross-pollen trans-fer.
The degree of reciprocity of stigma and anther heights in
the long- and short-styled morphs of eight Narcissus species with style-
If this evolutionary scenario is correct, an important
length dimorphism. For each morph, the stamen level that most closely
question that arises is why the transition to heterostyly is
corresponds in height to the stigma of the alternate morph is plotted. 1.
rare given the widespread occurrence of stigma-height
N. assoanus, 2. N. calcicola, 3. N. cuatrecasasii, 4. N. dubius, 5. N.
dimorphism in Narcissus. The only other well-docu-
gaditanus, 6. N. papyraceus, 7. N. rupicola, 8. N. albimarginatus. Data
mented case of heterostyly in the genus involves N. trian-
from a single population of each species were used in the comparison. drus, a tristylous species with occasional dimorphic pop-
The diagonal line represents complete reciprocity between stigma andanther heights. Note that both floral morphs of distylous N. albimargin-
ulations (Barrett et al., 1997). Narcissus albimarginatusatus (identified by arrows) fall on the line, whereas the remaining spe-
and N. triandrus, despite occurring in different sections
cies, all of which possess stigma-height dimorphism, are either above
of the genus, share a number of apparently convergent
or below the line. Source: 1–3, 5–7—Barrett, Lloyd, and Arroyo (1996);
floral characters suggesting a similar pollination syn-
4—Baker, Thompson, and Barrett (2000a); 8—this study.
drome. Both have pale-yellow pendulous flowers of sim-ilar size with a prominent corona. In addition, the fra-grance of both species is similar, at least as perceived by
been more intense in comparison with species possessing
us, and both flower during the same period. The major
pollinators of N. triandrus are large bees (Anthophora
Most distylous species possess a single stamen level
and Bombus), and it seems likely given their similarities
and therefore the assessment of the degree of reciprocal
in floral morphology that N. albimarginatus may also be
herkogamy in the floral morphs is straightforward. How-
adapted to bee pollination. The floral characteristics of
ever, dimorphic Narcissus species exhibit two stamen
both species are markedly different from Narcissus spe-
levels within a flower and therefore it is of interest to
cies with stigma-height dimorphism. These are mostly
compare the extent to which the two stamen levels cor-
adapted to hawk moths and butterflies and differ mainly
respond in height to stigmas of the opposite morph. In
in having smaller coronas with reduced depth and longer
N. albimarginatus one of the two stamen levels within a
floral tubes (Arroyo and Dafni, 1995; Barrett, Lloyd, and
flower corresponds closely in height, whereas the other
Arroyo, 1996; Baker, Thompson, and Barrett, 2000a, b).
is either above (long-styled morph) or slightly below
A shift of pollination system in Narcissus from lepidop-
(short-styled morph) stigmas of the alternate morph. This
teran to bee pollination could have provided the evolu-
pattern is also evident in Erythroxylum coca (Ganders,
tionary stimulus for the evolution of distyly from stigma-
1979b), an atypical distylous species with two stamen
height dimorphism. Elsewhere, Stone (1996) demonstrat-
levels. It is important to appreciate that there are often
ed that bees were more effective than lepidoptera at pro-
considerable disparities in the precise positions of anthers
and stigmas in heterostylous plants and the degree of re-
suerrensis, a distylous species with narrow-tubular co-
ciprocal herkogamy is rarely exact (Lloyd and Webb,
1992a). Nevertheless it seems likely that in N. albimar-
The evolution of a deep corona in association with the
ginatus, sex-organ positioning functions to promote sig-
shift to bee pollination may have been critical for the
nificant intermorph pollen transfer despite a lack of com-
evolution of heterostyly in Narcissus. The corona pro-
plete reciprocal herkogamy of all stamen levels. The 1:1
vides a longer vertical area for separation of the two sta-
morph ratio provides strong evidence that significant di-
men levels within a flower. This would enable more seg-
sassortative (intermorph) mating occurs in the population
regated pollen deposition on a pollinator’s body than is
as commonly occurs in distylous species.
possible in species with stigma-height dimorphism where
In accord with Lloyd and Webb’s (1992a, b) model of
anthers are closer together at the mouth of the narrow
the evolution of distyly we propose that distyly in N.
floral tube and packing constraints may restrict opportu-
albimarginatus is derived from an ancestral condition in-
nities for divergence in anther position (Barrett, Lloyd,
volving stigma-height dimorphism. First, on functional
and Arroyo, 1996). Deep coronas also occur elsewhere
grounds it would be difficult to imagine the selective
in Narcissus in the predominantly bee-pollinated sections
Bulbocodium and Pseudonarcissus, but species in these
lution and maintenance of stigma-height dimorphism in Narcissus:
sections are uniformly monomorphic for style length.
I. Floral variation and style-morph ratios. Heredity, In press.
Why then has heterostyly not developed in these groups?
———, ———, AND ———. 2000b. Evolution and maintenance of
stigma-height dimorphism in Narcissus: II. Fitness comparisons be-
We suggest it is the combination of a deep corona and a
tween style morphs. Heredity, In press.
long floral tube (absent from sections Bulbocodium and
BARRETT, S. C. H. [ED.]. 1992. Evolution and function of heterostyly.
Pseudonarcissus) that is necessary for the evolution of
heterostyly in Narcissus. These traits function to provide
———, W. W. COLE, J. ARROYO, M. B. CRUZAN, AND D. G. LLOYD.
more precise pollinator positioning and likely enable an-
1997. Sexual polymorphisms in Narcissus triandrus (Amaryllida-ceae): is this species tristylous? Heredity 78: 135–145.
ther height divergence and the segregated pollen transfer
———, L. K. JESSON, AND A. M. BAKER. 2000. Evolution of stylar
required for the origin and maintenance of the polymor-
polymorphisms in plants. Annals of Botany, In press.
———, D. G. LLOYD, AND J. ARROYO. 1996. Stylar polymorphisms
Because of its polyphyletic origins there is consider-
and the evolution of heterostyly in Narcissus (Amaryllidaceae). In
able variation in the heterostylous syndrome among an-
D. G. Lloyd and S. C. H. Barrett [eds.], Floral biology: studies onfloral evolution in animal-pollinated plants, 339–376. Chapman and
giosperm families and this has led to differing opinions
on how best to define the polymorphism (reviewed in
CHARLESWORTH, D., AND B. CHARLESWORTH. 1979. A model for the
Barrett, 1992). We believe that a morphological definition
evolution of distyly. American Naturalist 114: 467–498.
that concerns the relative positioning of sexual organs is
DULBERGER, R. 1964. Flower dimorphism and self-incompatibility in
the most useful perspective. In our opinion a population
Narcissus tazetta L. Evolution 18: 361–363.
FERNANDES, A. 1964. Contribution a` la connaissance de la ge´ne´tique
that contains two or three floral morphs possessing a re-
de l’he´te´rostylie chez le genre Narcissus. I. Re´sultats de quelques
ciprocal arrangement of stigma and anther heights qual-
croisements. Boletim da Sociedade Broteriana, Se´ries 2, 38: 81–
ifies a species as heterostylous regardless of whether it
possesses diallelic incompatibility or ancillary polymor-
GANDERS, F. R. 1979a. The biology of heterostyly. New Zealand Jour-
phisms of pollen and stigmas. This broader view empha-
———. 1979b. Heterostyly in Erythroxylum coca (Erythroxylaceae).
sizes the basic function of the polymorphism as a struc-
Botanical Journal of the Linnean Society 78: 11–20.
tural adaptation that increases the proficiency of polli-
˜es sobre algumas especies de Nar-
nator-mediated pollen transfer between plants. cissus, encontrados em Portugal. Boletim da Sociedade Broteriana
Lloyd and Webb (1992a, b) also emphasized functional
criteria in their treatment of the evolution of heterostyly
LOYD, D. G., AND C. J. WEBB. 1992a. The evolution of heterostyly. In S. C. H. Barrett [ed.], Evolution and function of heterostyly,
and proposed that distyly evolved from stigma-height di-
151–178. Springer-Verlag, Berlin, Germany.
morphism because of the influence of floral morphology
———, AND ———. 1992b. The selection of heterostyly. In S. C. H.
on pollen-transfer patterns. The occurrence of both stig-
Barrett [ed.], Evolution and function of heterostyly, 179–208.
ma-height dimorphism and distyly in Narcissus provides
KOHN, J. R., AND S. C. H. BARRETT. 1992. Experimental studies on the
evidence in support of their model. This association is of
functional significance of heterostyly. Evolution 46: 43–55.
particular significance because of the absence of diallelic
RICHARDS, A. J. 1997. Plant breeding systems. Chapman and Hall,
incompatibility in Narcissus (Dulberger, 1964; Sage et
al., 1999; Baker, Thompson, and Barrett, 2000b). While
———. 1998. Lethal linkage and its role in the evolution of plant
diallelic incompatibilty is generally a prerequisite for the
breeding systems. In S. J. Owens and P. J. Rudall [eds.], Repro-ductive biology in systematics, conservation and economic botany,
evolution of reciprocal herkogamy in some models for
71–83. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
the evolution of distyly (Charlesworth and Charlesworth,
MULLER-DOBLIES, D., AND U. MULLER-DOBLIES. 1989. Narcissus albi-
1979), those of Lloyd and Webb (1992a, b) are indepen-
marginatus, plate 1986. In The flowering plants of Africa, vol. 50,
dent of the type of compatibility system present in an-
part 2. Creda Press, Cape Town, South Africa.
cestral populations. Reciprocal herkogamy probably
SAGE, T. L., F. STRUMAS, W. W. COLE, AND S. C. H. BARRETT. 1999.
Differential ovule development following self- and cross-pollina-
evolved in N. albimarginatus to increase the effectiveness
tion in Narcissus triandrus (Amaryllidaceae). American Journal of
of pollen dispersal among plants because of altered pol-
lination conditions, perhaps associated with the isolated
STONE, J. L. 1996. Components of pollinator effectiveness in Psycho-
geographical distribution of this species. tria suerrensis, a tropical distylous shrub. Oecologia 107:504–512.
———, AND J. D. THOMSON. 1994. The evolution of distyly: pollen
transfer in artificial flowers. Evolution 48: 1595–1606.
VALDE´S, B. 1987. Narcissus. In B. Valde´s, S. Talavera, and E. Ferna´n-
dez-Galiano [eds.], Flora vascular de Andalucı´a Occidental, vol. 3,463–474. Ketres, Barcelona, Spain.
ARROYO, J., AND A. DAFNI. 1995. Variation in habitat, season, flower
WEBB, D. A. 1980. Narcissus. In T. G. Tutin, V. H. Heywood, N. A.
traits, and pollinators in dimorphic Narcissus tazetta L. (Amaryl-
Burges, D. M. Moore, D. H. Valentine, S. M. Walters, and D. A.
lidaceae) in Israel. New Phytologist 129: 135–145.
Webb [eds.], Flora Europaea, vol. 5, 78–84. Cambridge University
BAKER, A. M., J. D. THOMPSON, AND S. C. H. BARRETT. 2000a. Evo-
Bacterium background Rhizobia, is a col ective name of the genera Rhizobium, Sinorhizobium, Mesorhizobium and Bradyrhizobium , which are soil and rhizosphere bacteria of agronomic importance because they perform nitrogen-fixing symbioses with leguminous plants. Rhizobium and Sinorhizobium are in the family Rhizobiaceae, while Mesorhizobium and Bradyrhizobium members of Phyllobac
Conseil Scientifique du LOOF Recommandations concernant la l’amyloïdose rénale Mars 2007 Préambule : L'amyloïdose rénale est une maladie grave, assez rare, inéluctablement fatale, génétique mais dont le mode de transmission, à l'heure actuelle, est incomplètement connu. De ce fait, le gène muté n'étant pas encore – semble-t-il - isolé, un test diagnostic génétiqu
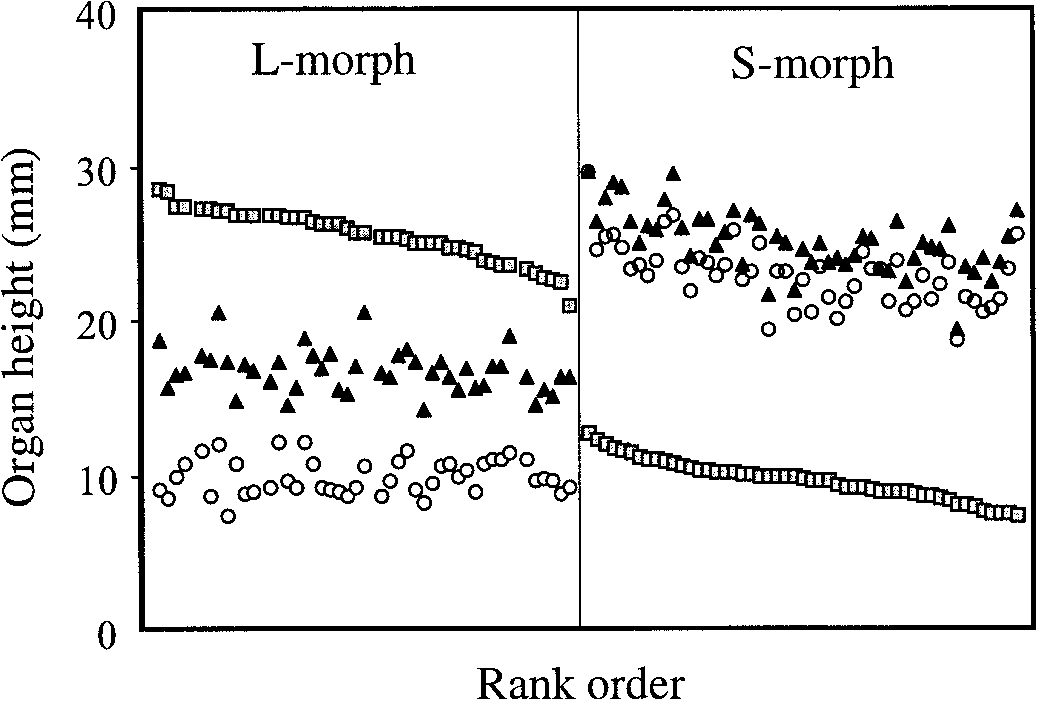 TABLE 1. Mean and (standard deviation) of stigma and anther heights
(mm) in the two floral morphs of distylous Narcissus albimargin-atus, a rare species from northwestern Morocco. See text for de-tails.
TABLE 1. Mean and (standard deviation) of stigma and anther heights
(mm) in the two floral morphs of distylous Narcissus albimargin-atus, a rare species from northwestern Morocco. See text for de-tails.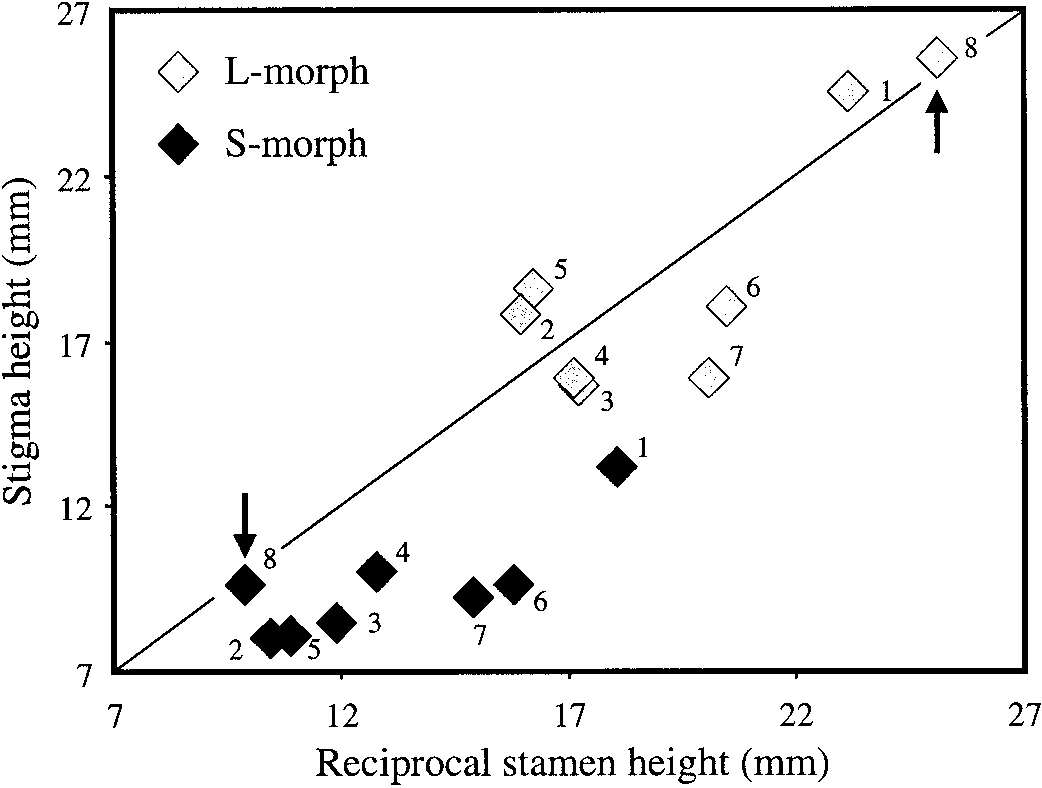 mechanisms that might favor the reverse evolutionary se-quence since this would likely reduce the proficiency ofcross-pollen dispersal between the floral morphs. Second,the widespread occurrence of stigma-height dimorphismin Narcissus, including species with varying degrees ofreciprocal herkogamy (Fig. 2), implies that variation ex-ists for the selection of distyly under the appropriate pol-lination environment. Finally, the isolated occurrence ofdistyly in a species apparently restricted to a single moun-tain in northwestern Morocco seems an unlikely place tolocate the ancestral polymorphic condition in Narcissus,unless of course this taxon represents a relictual speciesand all other distylous taxa have gone extinct. Clearly, aphylogeny of the genus would be required to fully resolvethese issues. In the meantime it seems to us that theweight of evidence favors the hypothesis that distyly inNarcissus is derived from stigma-height dimorphismthrough selection for more proficient cross-pollen trans-fer.
mechanisms that might favor the reverse evolutionary se-quence since this would likely reduce the proficiency ofcross-pollen dispersal between the floral morphs. Second,the widespread occurrence of stigma-height dimorphismin Narcissus, including species with varying degrees ofreciprocal herkogamy (Fig. 2), implies that variation ex-ists for the selection of distyly under the appropriate pol-lination environment. Finally, the isolated occurrence ofdistyly in a species apparently restricted to a single moun-tain in northwestern Morocco seems an unlikely place tolocate the ancestral polymorphic condition in Narcissus,unless of course this taxon represents a relictual speciesand all other distylous taxa have gone extinct. Clearly, aphylogeny of the genus would be required to fully resolvethese issues. In the meantime it seems to us that theweight of evidence favors the hypothesis that distyly inNarcissus is derived from stigma-height dimorphismthrough selection for more proficient cross-pollen trans-fer.