He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
The effects of estradiol on gonadotropin-releasing hormone neurons in the developing mouse brain
General and Comparative Endocrinology 112, 356–363 (1998) Article No. GC987134 The Effects of Estradiol on Gonadotropin-Releasing Hormone Neurons in the Developing Mouse Brain Matthew S. Grober,1 Greg M. Winterstein, Asif A. Ghazanfar,2 and Victor P. Eroschenko* Department of Biological Sciences, *WAMI Medical Program, University of Idaho, Moscow, Idaho 83844-3051The hypothalamic–pituitary–gonadal (HPG) axis plays a
The perinatal organization of the vertebrate brain
critical role in the control of reproduction. Two key
plays a major role in determining adult reproductive
hormonal components of the HPG axis are gonadal
function. Two brain regions that are involved in the
steroids and gonadotropin-releasing hormone (GnRH).
control of reproductive behavior and physiology are
Gonadal steroids are known to organize the development
the hypothalamus and the neurohypophysis, which, in
of neural substrates which control adult reproductive
conjunction with the gonads, constitute the hypotha-
behavior; GnRH is required for normal reproductive
lamic–pituitary–gonadal axis (HPG axis). Two impor-
structure and function. The possibility that gonadal
tant hormonal components of the HPG axis are go-
steroids may produce organizational changes in the
nadal steroids and gonadotropin-releasing hormone
pattern of GnRH staining observed in the brain is investigated through the use of injections of estradiol to
Gonadal steroids mediate developmental organiza-
neonatal mice and subsequent GnRH immunocytochem-
tion of the neural and nonneural substrates that are
istry at 2 months of age. Our results indicate that the
critical for adult reproductive function (Phoenix et al.,number of GnRH-immunoreactive (GnRH-ir) cells is
1959). Experimental manipulation of testosterone re-
normally lower in females than males. Estradiol did not
sults in changes in the expression of male-like external
affect the number of GnRH-ir cells in females, but
genitalia and frequency of mounting behavior by
significantly increased the number of GnRH-ir cells in
female offspring (Phoenix et al., 1959), the number of
males, suggesting that early exposure to estradiol results
neurons in the spinal nucleus of the bulbocavernosus
in masculinization of the GnRH axis of males.
in both males and females (Breedlove and Arnold,
1983), and anogenital distance (AGD) (Clemens et al.,Key Words: GnRH; sexual differentiation; hypothalam-
1978). The sexually dimorphic nucleus of the preoptic
ic–preoptic area; neonates; estradiol; immunocytochem-
area (SDN-POA) is one of several regions of the rodent
istry; mouse; organization.
brain that is influenced during development by circu-lating gonadal steroids (Gorski et al., 1978). The SDN-POA is larger in males than females (Gorski et al.,1980), and injection with testosterone in early life
1To whom correspondence should be addressed at: Department of
causes this region to increase in size in both gonadecto-
Life Sciences, Arizona State University West, P.O. Box 37100, 4701
mized males and normal females (Rhees et al., 1990).
West Thunderbird Avenue, Phoenix, AZ 85069–7100. Fax: (602)
The estrogen antagonist tamoxifen and the androgen
543–6073. E-mail: [email protected].
2Present address: Department of Neurobiology, Box 3209, Duke
antagonist cyproterone acetate have been used to show
University Medical Center, Durham, NC 27710.
the estrogenic mediation of sexual dimorphism in the
All rights of reproduction in any form reserved.
SDN-POA (Do¨hler et al., 1984, 1986). More recently,
ever, this analysis was qualitative and thus did not
antisense oligonucleotides which block the production
examine quantitative differences in the effect of estra-
of the estrogen receptor have been used to show that in
diol on the number of GnRH cells. Data from radioim-
female rats SDN-POA size and parastrial nuclear size
munoassays on brain tissue from neonatally estro-
are mediated by estrogen and that estrogen influences
genized rats suggest that early estrogen treatment
the development of both lordosis and open field
modifies the hypothalamic mechanism involved in the
behavior (McCarthy et al., 1993). These data suggest
release of LHRH (Hayashi et al., 1991). Two previous
that aromatization of testosterone to estradiol is respon-
studies have quantitatively examined GnRH cells in
sible for several key components of the masculiniza-
mice (Hoffman and Finch, 1986; Wray et al., 1989).
tion process. Thus, gonadal steroids have dramatic
Wray et al. (1989) examined the progenitor cells that
organizational effects on areas of the hypothalamus
give rise to forebrain GnRH cells and provided counts
and preoptic area that play a key role in the control of
of GnRH cells from embryonic day 10.5 through
reproduction. It is possible that the organizational
adulthood. Hoffman and Finch (1986) looked at GnRH
effects of gonadal steroids may also influence the
cells during aging in a different strain of mice and
development of the hypothalamic nuclei responsible
found that onset of reproductive dysfunction did not
correlate with a loss of GnRH forebrain cells. Presently,
The gonadotropin-releasing hormones are a family
no data are available on the effects of estradiol on the
of decapeptide hormones found in all vertebrates
development of the GnRH axis in mice. This study
examined thus far and which represents an important
tests the hypothesis that early postnatal estradiol
link between the brain and reproduction. In rodents,
treatment affects the number of GnRH-immunoreac-
the majority of the GnRH-producing cells are in the
tive (GnRH-ir) neurons in the preoptic area of mice of
preoptic region of the hypothalamus (reviewed in
both sexes during the first 2 months of development.
Silverman et al., 1994). Most of the GnRH cells arelocated amidst the diagonal band of Broca, the bednucleus of the stria terminalis, the preoptic region(periventricular, medial, and lateral), and the anterior
MATERIALS AND METHODS
hypothalamus (Silverman et al., 1994). The majority ofGnRH cells located in the preoptic region project to the
Animals
median eminence and are associated with the hypophy-seal portal system (Silverman et al., 1994). Gonadotro-
Adult mice of ND4 Swiss Webster strain were
pin-releasing hormone is primarily responsible for
purchased from Simonsen Labs (Gilroy, CA), given
causing the release of luteinizing hormone and follicle-
food and water ad libitum, kept on a 12-h light–12-h
stimulating hormone from the anterior pituitary. It is
dark regimen, and allowed to breed. Within 24 h after
also involved in the activation of lordosis in the female
birth, the dams with their litters (housed together)
(Moser and Mathiesen, 1996; Pfaff, 1973; Sakuma and
were randomly assigned to different experimental
Pfaff, 1980). Moreover, in both mice and humans
groups. Immediately after group assignment, neonatal
(reviewed in Silverman et al., 1994), improper develop-
pups received the first of 14 daily intraperitoneal
ment and or molecular regulation of the GnRH fore-
injections (0.05 ml) of either sesame oil (control) or 10.0
brain nuclei results in hypogonadism and subsequent
mg of 17-estradiol in sesame oil. All injections were
infertility. Thus, early and proper development of the
administered with a 27-gauge needle using filtered
forebrain GnRH axis is a necessary requirement for
sesame oil as a vehicle. The choice of estradiol dosage
normal adult reproductive function in several mamma-
was based on previous experiments where this dose
induced significant morphological and biochemical
Neonatal treatment of rats with estradiol decreases
alterations in the reproductive organs of immature
the number of cells stained for GnRH in the male and
female mice (Eroschenko et al., 1995). We were notified
increases the number of cells in the female within the
as to the availability of brain tissue from these animals
first 10 days of life (Elkind-Hirsch et al., 1981). How-
after the study was completed, and the established
All rights of reproduction in any form reserved.
experimental design places some limitations on our
cific neuroanatomical landmarks including the merger
of the right and left radiations of the frontal aspect of
At 2 months of age (56–60 days), all treated mice
the corpus callosum, anteriorly, to the appearance of
were anesthetized, injected with 100 ml heparin (1000
the fornix and the disappearance of the anterior com-
units/ml), and perfused with 0.9% sodium chloride
missure, posteriorly. This forebrain area includes the
and then with 4% paraformaldehyde in 0.1 M phos-
medial and lateral preoptic areas, the medial and
phate-buffered saline. Anogenital distance and body
lateral septal nuclei, and the diagonal band of Broca.
weight were recorded before perfusion, whereas ovar-
Total cell counts in the forebrain for each animal were
ian weight was recorded after perfusion (testes weight
tabulated. The GnRH-ir cells in some brain sections
was not recorded). The brains were removed, blocked,
were not visible because of poor staining. It is not clear
sunk in 30% sucrose overnight at 4° C for cryoprotec-
why some sections did not stain well, but poorly
tion, sectioned at 50 µm in the coronal plane on a
stained sections were distributed haphazardly among
cryostat, and then stored in 0.1 M phosphate buffer
brains and no brains were composed entirely of poorly
(PB) at 4°C until the immunostaining was performed.
stained sections. Thus, GnRH-ir cell number in brainswith poorly stained sections would not be a reliableindicator of the total number of cells in those brains. In
Immunocytochemistry
order to exclude brains with poorly stained sections,
Immunocytochemistry was carried out on free-floating
group averages were calculated from the four highest
sections using culture plates. Briefly, the sections were
values in each group. The top four brains were chosen
rinsed twice in PB with 0.4% Triton X-100 (PBX) for 5
for two reasons: (1) one treatment group only had four
min. The sections were then incubated for 1 h in the
brains that showed robust staining for all sections and
presoak solution (3% normal goat serum in PBX) at
(2) to maintain equal sample sizes across all treat-
room temperature on a shaker. The sections were then
ments. This ensured that the staining quality was
incubated with the primary antibody (monoclonal
robust in all brains used in the analyses (Fig. 1).
antibody to GnRH, LR 132 (Park and Wakabayshi,
Differences between the means for all groups were
1986), diluted 1:1000 in the presoak solution) overnight
analyzed using analysis of variance (ANOVA). The
(at least 16 h) at room temperature on a shaker. The
statistical significance of the differences among the
sections were rinsed twice in PBX for 5 min. Biotinyl-
individual groups was analyzed using Fisher’s LSD.
ated secondary antibody (Kirkegaard & Perry Labora-
Linear regression was used to evaluate the relationship
tories, Gaithersburg, MD; KPL) was then applied for 1
between gonad size and body size (StatView 4.01 for
h at room temperature on a shaker. The sections were
the Macintosh, Abacus Concepts, Inc.). All data are
rinsed twice with PBX for 5 min. Incubation of the
presented as the mean Ϯ standard error of the mean.
sections with streptavidin–peroxidase (KPL) was at roomtemperature for 1 h. The sections were rinsed twicewith PBX for 5 min. DAB (diaminobenzidine), diluted
according to the manufacturer’s instructions (KPL),was applied to the sections for 10 min. The sectionswere rinsed in PB to stop the DAB reaction, mounted
Localization of GnRH-Producing Neurons
on slides, air dried, dehydrated, and coverslipped.
Our identification of GnRH-ir cells in the mouse
forebrain (Fig. 1) is similar to previously published
Cell Count Analysis
work (reviewed in Silverman et al., 1994). All GnRH-irneurons were located in forebrain regions that were
We counted all stained cells in which a nucleus and
included in the quantitative analyses.
one or more cell processes were visible. We includedthe second criterion to avoid double counting cells that
Cell Counts
were cut through the plane of the nucleus and thuswould exhibit a nucleus on more than one section. The
There were significant differences in the number of
forebrain region examined was delineated using spe-
GnRH-ir cells among the four treatment groups (Fig. 2;
All rights of reproduction in any form reserved.
Characteristics of the mouse GnRH forebrain system. (A) Cross-section of the mouse brain. CC, corpus callosum; AC, anterior
commissure; ON, optic nerve. Bar, 700 µm. The box indicates the area of B (not to scale). (B) A photomicrograph of representative GnRH-irneurons in the preoptic area of the mouse brain at 2 months of age. Bar, 100 µm. Arrow, GnRH-ir neurons shown in C. (C) Higher magnificationphotomicrograph of representative GnRH-ir neurons. Bar, 10 µm.
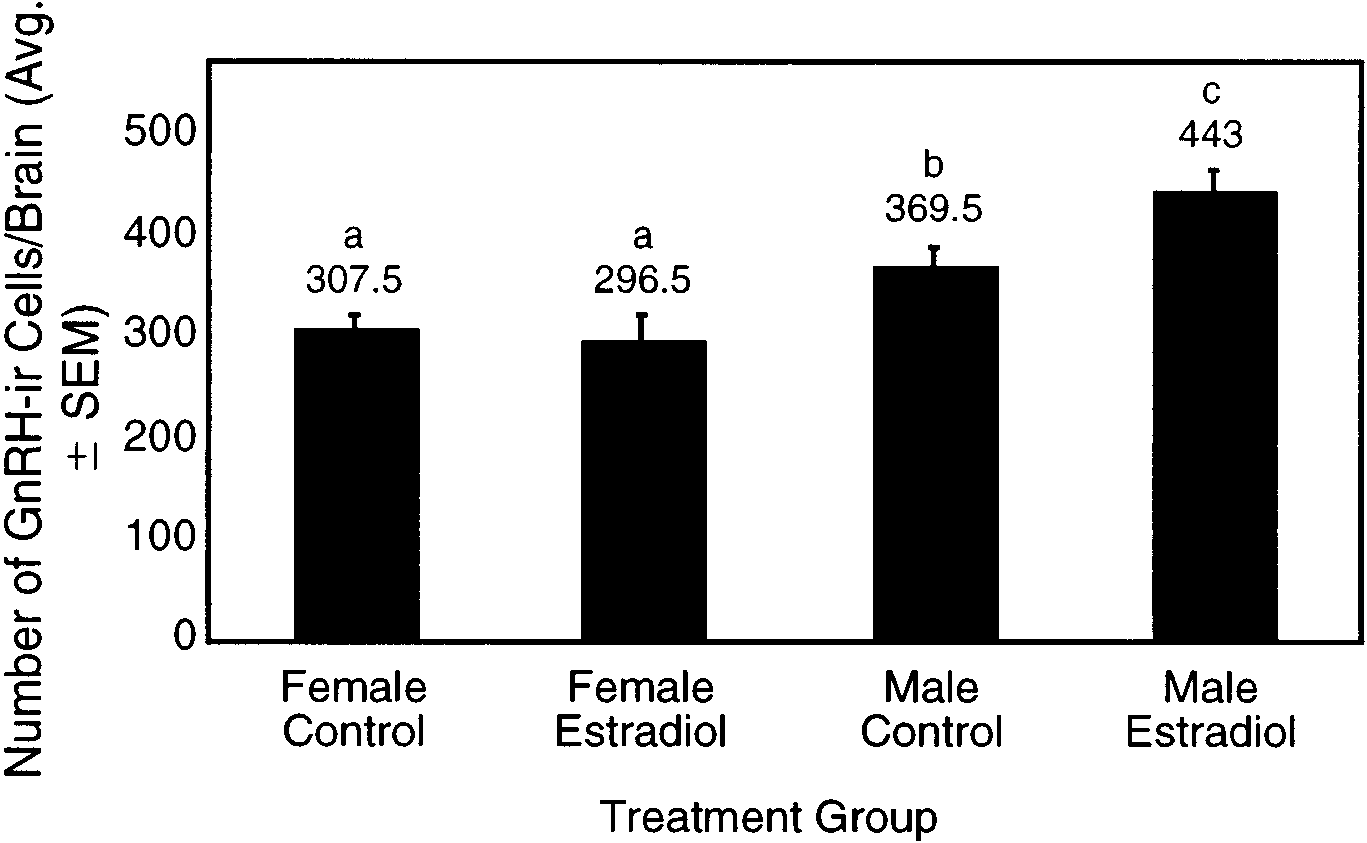
ANOVA, F ϭ 10.47, df ϭ 3, P Ͻ 0.01). The number of
(369.5 Ϯ 16.56). Similarly, estradiol-treated males exhib-
GnRH-ir cells in control males was 17% greater than in
ited a significantly greater number of cells than estra-
control females (statistically significant at P ϭ 0.057,
diol-treated females (P Ͻ 0.001).
LSD). There were no significant differences in thenumber of GnRH cells between control females (307.5 Ϯ
Somatic Characteristics
13.88 cells per brain) and estradiol-treated females(296.5 Ϯ 28.37; P ϭ 0.72) (Fig. 2). In contrast, estradiol-
The body weight of the female mice was signifi-
treated males have significantly more GnRH-ir neu-
cantly increased by estradiol treatment (P Ͻ 0.01, Table
rons (443.0 Ϯ 21.53; P ϭ 0.03) than control males
1). The male controls were, as expected, heavier than
The effects of estradiol treatment on GnRH immunoreactivity in male and female mouse brains at 2 months of age. The columns and
error bars represent the means and standard errors of the number of GnRH-ir neurons per brain (n ϭ 4), respectively. Different letters (a, b, c)indicate significant differences between the groups as determined using Fisher’s LSD test (P Ͻ 0.05).
All rights of reproduction in any form reserved.
Effects of Estradiol on Morphological Characteristics of Two-Month-Old Mice
*Significant difference (P Ͻ 0.05) between control and estradiol treated mice. For all cells in the table, n ϭ 4.
the female controls (P Ͻ 0.001, Table 1). Anogenital
is no statistically significant relationship. Thus, our
distance was not affected by estradiol treatment in
data suggest a sexual dimorphism in the number of
either males or females (Table 1). However, the AGD
GnRH-ir neurons in the preoptic area of the mouse
data show the expected sexual dimorphism and are in
brain. This pattern resembles observations in the rat,
agreement with previously published values showing
where males were reported to have a greater number
males to have a greater AGD than females (Simon and
of GnRH-ir neurons than females; however, this differ-
Cologer-Clifford, 1991). Ovarian weight showed a
ence was not quantified (Elkind-Hirsch et al., 1981). A
nonsignificant decrease in estradiol-treated females
second study on rats did not find a difference in the
relative to control females (P ϭ 0.16, Table 1); however,
number of GnRH cells between males and females, but
only estradiol-treated females had cystic ovaries.
this study did not indicate the estrous state of thefemale animals (see discussion below) (Silverman etal., 1994).
Our data differ from two previous studies on mice in
DISCUSSION
several ways. We counted fewer cells (300–400) com-pared to total GnRH cell numbers in Wray et al. (1989)
Gonadal steroids affect the development and sexual
(about 800) and Hoffman and Finch (1986) (about 650).
differentiation of all vertebrates that have been exam-
While this is a dramatic difference, several sources of
ined. Our results indicate that the number of preoptic
variation may account for these differences. All three
GnRH-ir cells in adult mice may be dependent upon
studies used sections of different thickness (present
the steroidal milieu during development. Since several
study, 50µm; Wray et al., 12–14 µm; Hoffman and
brain areas, including the suprachiasmatic nucleus, are
Finch, 30µm) and employed different criteria for count-
not affected by estrogens (Do¨hler et al., 1986), it is likely
ing cells (we required a nucleus and at least one
that the changes we observed in the number of
neurite, Hoffman and Finch required a nucleus, and
GnRH-ir cells are not a result of nonspecific effects of
Wray et al. did not provide any specific criteria). The
use of thinner sections and less conservative criteria
The comparison between untreated males and fe-
for cell counting would have increased the probability
males suggests that the number of GnRH-ir neurons in
of multiply counting cells, resulting in the higher cell
mice may be sexually dimorphic. One explanation for
our result is that gonadal steroids present in the
Each of the studies utilized different strains of mice
developing male and female differ, resulting in differ-
which have been genetically isolated for many genera-
ent numbers of GnRH-ir neurons at 2 months of age.
tions. Given that normal reproductive function can be
An alternative explanation is that because male mice
maintained with a very small number of GnRH cells
are on average larger than female mice (Table 1), they
(Silverman et al., 1994), there is no reason to expect
may have a larger brain and hence a larger number of
different strains to maintain similar cell numbers. An
GnRH-ir neurons. However, regression analysis be-
additional difference between the studies involves
tween body weight and the number of GnRH-ir
gonadal influences. Hoffman and Finch ovariecto-
neurons, for the animals in our study, shows that there
mized their mice, while Wray et al. did not. We were
All rights of reproduction in any form reserved.
unable to include gonadectomized controls in this
change in the AGD indicating no effects on androgen-
study. However, Hoffman and Finch found that ovari-
mediated characteristics. The increase in the number of
ectomy did not significantly affect cell numbers in
GnRH-ir neurons contrasts with prior data from the rat
which showed a decrease in the number of GnRH-ir
There are a number of factors that can affect the
cells after treatment with estradiol in male rats (Elkind-
amount of GnRH immunostaining observed in adult
Hirsch et al., 1981). However, our data were collected at
rodent brains. Changing gonadal hormone titers over
2 months of age while the data from the rat were
the estrous cycle of the rat are correlated with GnRH
collected before 10 days of age. After neonatal treat-
content of the hypothalamus (Araki et al., 1975; Free-
ment with estradiol, hypothalamic concentrations of
man, 1988), and GnRH mRNA expression increases in
GnRH are lower in early neonatal life (5–7 days) and
late proestrus (Silverman et al., 1994). The number of
higher later in development (60 days) when compared
GnRH neurons that are stained in the brain of the rat is
with those of the untreated control rat (Goomer et al.,
significantly lower in late estrus compared with the
1977). This could explain the differences between our
remainder of the cycle (Ronnekleiv and Kelly, 1986;
results and those of Elkind-Hirsch et al. (1981).
Silverman and Witkin, 1994), although at least one
Since the male mouse brain contains active aro-
study has shown that the number of neurons stained
matase (Wozniak et al., 1992), it is possible that testos-
for GnRH may not change during the estrous cycle of
terone in normal males is aromatized to estradiol
the rat (Marks et al., 1993). Low estradiol levels reduce
causing an increase in the number of GnRH-ir neurons
the release of GnRH in the female rat (Silverman et al.,
above that of normal females. This idea is consistent
1994), while a surge of estradiol precipitates the release
with our data (and prior data, Elkind-Hirsch et al.,
of GnRH and subsequently the release of luteinizing
1981) showing a greater number of GnRH-ir neurons
hormone from the pituitary (Kaynard et al., 1988; Krey
in males. Taken together, these results suggest that this
and Parsons, 1982). Thus, variation in GnRH cell
sexual dimorphism may be mediated by the aromatiza-
number may, or may not, occur over the course of the
tion of testosterone to estradiol. However, it is known
estrous cycle. Since the estradiol-treated female mice in
that both 5␣-dihydrotestosterone and estradiol can
our study were presumed to be in a state of persistent
affect the number of GnRH-ir neurons in rats (Silver-
vaginal estrus or diestrus, this variation should have a
man et al., 1994) and frogs (Iela et al., 1994). Thus, it is
minimal effect on the variation in GnRH cell number
probably unlikely that aromatase alone is responsible
for the difference in GnRH-ir neuron numbers.
Neonatal treatment with estradiol affects the uterus
The exact mechanisms whereby estradiol mediates
by causing a decrease in the number of estrogen
GnRH cell function in the mammal brain are not
receptors in the 2-month-old rat (Csaba and Incezefi-
entirely clear. Several studies indicate that GnRH-
Gonda, 1992). A similar effect may occur in the brain,
producing neurons in a variety of mammalian species
decreasing the estrogen receptor-binding capacity and
do not possess estrogen receptors as adults (Herbison
hence decreasing the adult sensitivity of GnRH neu-
and Theodosis, 1992; Lehman and Karsch, 1993; Silver-
rons to estradiol. Thus estradiol may have an activa-
man et al., 1994). To our knowledge, there are no
tional effect, but this would be masked by the de-
existing studies that examine potential colocalization
creased sensitivity of these neurons. Unfortunately, the
of estrogen receptors in GnRH cells of preadult mam-
design of this study prevents the testing of this hypoth-
mals. Given that the GnRH gene in mice (Radovick et
esis. Regardless, the lack of difference between the
al., 1992) contains a region that binds the estrogen
control and estradiol-treated females indicates that
receptor (estrogen response elements), it is possible
neonatal estradiol treatment does not appear to have
that GnRH-producing neurons do express the estrogen
an effect on the number of GnRH-producing neurons
receptor during a specific developmental period. Tran-
sient expression of estrogen receptors during develop-
Males treated with estradiol show a significant
ment has been shown in the rat cortex (Yokosuka et al.,
increase in the number of GnRH cells compared to
1995). Although the estrogen receptor has not been
control males. At the same time, our data show no
identified in vivo in adult GnRH cells, there are cell
All rights of reproduction in any form reserved.
lines that do produce GnRH and may express the
REFERENCES
estrogen receptor (Poletti et al., 1994). An alternativehypothesis is that a more complex mechanism isinvolved that does not require the GnRH-producing
Aihara, M., and Hayashi, S. (1989). Induction of persistent diestrus
followed by persistent estrus is indicative of delayed maturation
neurons to respond directly to estrogen (Silverman et
of tonic gonadotropin-releasing systems in rats. Biol. Reprod. 40(1),
While our data do not show a significant decrease in
Araki, S., Ferin, M., Zimmerman, E. A., and Wiele, R. L. V. (1975).
ovarian weight in estradiol-treated females (Table 1)
Ovarian modulation of immunoreactive gonadotropin-releasing
this trend is consistent with earlier data in the adult rat
hormone (GnRH) in the rat brain: Evidence for a differential effect
(Brawer et al., 1986; Kikuyama, 1962) and mouse
on the anterior and mid-hypothalamus. Endocrinology 96(3), 644– 650.
(Eroschenko et al., 1995; Martinez and Swartz, 1991).
Brawer, J., Munoz, M., and Farookhi, R. (1986). Development of the
Only the estradiol-treated animals in our study had
polycystic ovarian condition (PCO) in the estradiol valerate-
cystic ovaries, which is consistent with a state of
treated rat. Biol. Reprod. 35, 647–655.
estradiol-induced persistent vaginal estrus or diestrus
Breedlove, S. M., and Arnold, A. P. (1983). Hormonal control of a
(Aihara and Hayashi, 1989; Kikuyama, 1962), as shown
developing neuromuscular system. I. Complete demasculiniza-tion of the male rat spinal nucleus of the bulbocavernosus using
in earlier studies (Aihara and Hayashi, 1989; Walters,
the anti-androgen flutamide. J. Neurosci. 3, 417–423.
Clemens, L., Glaude, B., and Coniglio, L. (1978). Prenatal endog-
In conclusion, we have shown that male mice have
enous androgenic influences on masculine sexual behavior and
more preoptic GnRH-ir neurons than females, and
genital morphology in male and female rats. Horm. Behav. 10,
estradiol treatment of neonatal mice causes sex-
specific increases in the number of GnRH-ir neurons;
Csaba, G., and Incezefi-Gonda, A. (1992). Life-long effect of a single
neonatal treatment with estradiol or progesterone on rat uterine
males respond and females do not. This is consistent
estrogen receptor binding capacity. Horm. Metab. Res. 24, 167–171.
with the notion of sexually dimorphic neural sub-
Do¨hler, K., Coquelin, A., Davis, F., Hines, M., Shryne, J., Sickmoller,
strates developing at an early age. In response to high
P., Jarzab, B., and Gorski, R. (1986). Pre- and postnatal influence of
levels of exogenous estradiol, males show an increase
an estrogen antagonist and an androgen antagonist on differentia-
in GnRH-producing neurons in the brain. It is possible
tion of the sexually dimorphic nucleus of the preoptic area in male
that these changes in the number of GnRH-ir neurons
and female rats. Neuroendocrinology 42, 443–448.
Do¨hler, K., Srivastava, S., Shryne, J., Jarzab, B., Sipos, A., and Gorski,
are normally mediated through the aromatization of
R. (1984). Differentiation of the sexually dimorphic nucleus in the
testosterone to estradiol in the brain. Finally, it is
preoptic area of the rat brain is inhibited by postnatal treatment
known that by changing the neonatal steroidal milieu,
with an estrogen antagonist. Neuroendocrinology 38, 297–301.
the organization of the brain can be altered in ways
Elkind-Hirsch, K., King, J. C., Gerall, A. A., and Arimura, A. (1981).
that have a significant impact on adult reproduction.
The luteinizing hormone-releasing hormone (LHRH) system in normal and estrogenized neonatal rats. Brain Res. Bull. 7, 645–654.
In summary, one interpretation, consistent with our
Eroschenko, V. P., Abuel-Atta, A. A., and Grober, M. S. (1995).
results, is an organizational effect of estradiol on the
Neonatal exposures to technical methoxychlor alters ovaries in
number of GnRH-ir neurons in the male mouse. A
adult mice. Reprod. Toxicol. 9, 379–387.
putative mechanism for this effect is suggested by the
Freeman, M. (1988). The ovarian cycle of the rat. In ‘‘The Physiology
higher number of cells in estradiol-treated males.
of Reproduction’’ (E. Knobil, J. Neill, and L. Ewing, Eds.), pp. 1893–1928. Raven Press, New York.
Goomer, N., Saxena, R., and Sheth, A. (1977). Effect of neonatal
testosterone and oestradiol treatment on the development of thehypothalamo–hypophysial axis in the female rat. J. Reprod. Fertil.ACKNOWLEDGMENTS 50, 239–243.
Gorski, R., Gordon, J., Shryne, J., and Southam, A. (1978). Evidence
for a morphological sex difference within the medial preoptic area
The authors thank Kelly Lane for obtaining body weight and
of the rat brain. Brain Res. 148, 333–346.
anogenital distance measurements, Laurie Kawabata for the ovarian
Gorski, R., Harlan, R., Jacobson, C., Shryne, J., and Southam, A.
weights, Jason Miranda for editorial assistance, and Mark DeSantis,
(1980). Evidence for the existence of a sexually dimorphic nucleus
Rodney Mead, and Catherine Ulibarri for comments on earlier drafts
in the preoptic area of the rat. J. Comp. Neurol. 193, 529–539.
of the manuscript. This research was supported by an NIH IDeA
Hayashi, S., Aihara, M., and Wakabayashi, K. (1991). Content and
grant and a University of Idaho Seed Grant to M.S.G.
distribution pattern of luteinizing hormone-releasing hormone
All rights of reproduction in any form reserved.
(LHRH) in the hypothalamus of neonatally estrogenized female
nate on the tissues mediating mating behavior in the female
rats. Neurosci. Res. 12(2), 366–378.
guinea pig. Endocrinology 65, 369–382.
Herbison, A. E., and Theodosis, D. T. (1992). Localization of oestro-
Poletti, A., Melcangi, R. C., Negri-Cesi, P., Maggi, R., and Martini, L.
gen receptors in preoptic neurons containing neurotensin but not
(1994). Steroid binding and metabolism in the luetinizing hormone-
tyrosine hydroxylase, cholecystokinin or luteinizing hormone-
releasing hormone-producing neuronal cell line GT1–1. Endocrinol-
releasing hormone in the male and female rat. Neuroscience 50(2), ogy 135(6), 2623–2628.
Radovick, S., Wondisford, F., Wray, S., Ticknor, C., Nakayama, Y.,
Hoffman, G., and Finch, C. E. (1986). LHRH neurons in the female
Cutler, G. J., Weintraub, B., Westphal, H., and Lee, E. (1992).
C57BL/6J mouse brain during reproductive aging: No loss up to
Characterization, expression, and estradiol regulation of the hu-
middle age. Neurobiol. Aging 7, 45–48.
man GnRH gene. In ‘‘Modes of Action of GnRH and GnRH
Iela, L., D’Aniello, B., Di-Meglio, M., and Rastogi, R. (1994). Influ-
Analogs’’ (W. J. Crowley and P. Conn, Eds.), pp. 85–105. Springer-
ence of gonadectomy and steroid hormone replacement therapy
on the gonadotropin-releasing hormone neuronal system in the
Rhees, R., Shryne, J., and Gorski, R. (1990). Termination of the
anterior preoptic area of the frog (Rana esculenta) brain. Gen.
hormone-sensitive period for differentiation of the sexually dimor-
Comp. Endocrinol. 95, 422–431.
phic nucleus of the preoptic area in male and female rats. Brain
Kaynard, A., Malpaux, B., Robinson, J., Wayne, N., and Karsch, F. Res. Dev. Brain Res. 52, 17–23.
(1988). Importance of pituitary and neural actions of estradiol in
Ronnekleiv, O., and Kelly, M. (1986). Luteinizing hormone-releasing
induction of the luteinizing hormone surge in the ewe. Neuroendo-
hormone neuronal system during the estrous cycle of the female
crinology 48, 296–303.
rat: effects of surgically induced persistent estrus. Neuroendocrinol-
Kikuyama, S. (1962). Inhibition of induction of persistent estrus by
ogy 43, 564–576.
chloropromazine in the rat. Annot Zool. Jpn. 95, 147–154.
Sakuma, Y., and Pfaff, D. (1980). LH-RH in the mesencephalic central
Krey, L., and Parsons, B. (1982). Characterization of estrogen stimuli
grey can potentiate lordosis reflex of female rats. Nature 283,
sufficient to initiate cyclic luteinizing hormone release in acutely
ovariectomized rats. Neuroendocrinology 34, 315–322.
Silverman, A.-J., and Witkin, J. W. (1994). Biosynthesis of gonadotro-
Lehman, M. N., and Karsch, F. J. (1993). Do gonadotropin-releasing
pin-releasing hormone during the rat estrous cycle: A cellular
hormone, tyrosine hydroxlase-, and B-endorphin-immunoreactive
analysis. Neuroendocrinology 59, 545–551.
neurons contain estrogen receptors? A double-label immunocyto-
Silverman, A. J., Livne, I., and Witkin, J. (1994). The gonadotropin-
chemical study in the suffolk eve. Endocrinology 133(2), 887–895.
releasing hormone (GnRH) neuronal systems: Immunocytochem-
Marks, D. L., Smith, M. S., Vrontakis, M., Clifton, D. K., and Steiner,
istry and in situ hybridization. In ‘‘The Physiology of Reproduc-
R. A. (1993). Regulation of galanin gene expression in gonadotro-pin-releasing hormone neurons during the estrous cycle of the rat.
tion’’ (E. Knobil and J. Neill, Eds.), Vol. 1, pp. 1683–1710. Raven
Endocrinology 132(4), 1836–1844.
Martinez, E., and Swartz, W. (1991). Effects of methoxychlor on the
Simon, N. G., and Cologer-Clifford, A. (1991). In utero contiguity to
reproductive system of the adult female mouse. 1. Gross and
males does not influence morphology, behavioral sensitivity to
histological observations. Reprod. Toxicol. 5, 139–147.
testosterone, or hypothalamic androgen binding in CF-1 female
McCarthy, M. M., Schlenker, E. H., and Pfaff, D. W. (1993). Enduring
mice. Horm. Behav. 25, 518–530.
consequences of neonatal treatment with antisense oligodeoxy-
Walters, L. M., Rourke, A. W., and Eroschenko, V. P. (1993). Purified
nucleotides to estrogen receptor messenger ribonucleic acid on
methoxychlor stimulates the reproductive tract in immature fe-
sexual differentiation of rat brain. Endocrinology 133(2), 433–439.
male mice . Reprod. Toxicol. 7, 599–606.
Moser, E. I., and Mathiesen, L. I. (1996). Relationship between
Wozniak, A., Hutchison, R., and, Hutchison, J. (1992). Localization of
neuronal activity and brain temperature in rats. NeuroReport 7(11),
aromatase activity in androgen target areas of the mouse brain. Neurosci. Lett. 146, 191–194.
Park, M. K., and Wakabayshi, K. (1986). Preparation of a monoclonal
Wray, S., Philip, G., and Gainer, H. (1989). Evidence that cells
antibody to common amino acid sequence of LHRH and its
expressing luteinizing hormone-releasing hormone mRNA in the
application. Endocrinol. Jpn. 33, 257–272.
mouse are derived from progenitor cells in the olfactory placode.
Pfaff, D. (1973). Luteinizing hormone-releasing factor potentiates
Proc Natl. Acad. Sci. USA 86, 8132–8136.
lordosis behavior in hypophysectomized ovariectomized female
Yokosuka, M., Okamura, H., and Hayashi, S. (1995). Transient
rats. Science 182, 1148–1149.
expression of estrogen receptor-immunoreactivity (ER-IR) in the
Phoenix, C. H., Goy, R. W., Gerall, A. A., and Young, W. C. (1959).
layer V of the developing rat cerebral cortex. Brain Res. 84(1),
Organizing action of prenatally administered testosterone propio-
All rights of reproduction in any form reserved.
ÖKOLOG SOMMERAKADEMIE 2010 IN VIKTORSBERG Dramatechnik Erzählerbericht Die Geschichte, um die es geht, handelt von Gerechtigkeit und fehlender Gerechtigkeit. Sie handelt von einem indianischen Stamm und von reichen westlichen Pharmakonzernen, das sind große Firmen, die mit Medikamenten, Nahrungsmittelzusätzen, Heilmitteln und Schönheitsprodukten ihr Geld verdienen. Die Geschichte
HKSAR’s unique advantages in Pan-Pearl River Delta Region I attended the Pan-Pearl River Delta Regional Co-operation and Development Forum (PPRD Forum) in Hainan on November 30 and December 1. It was the first time I attended the PPRD Forum and also my first major visit outside Hong Kong since I took office as Chief Executive. The trip proved fruitful. In particular, we achieved encoura
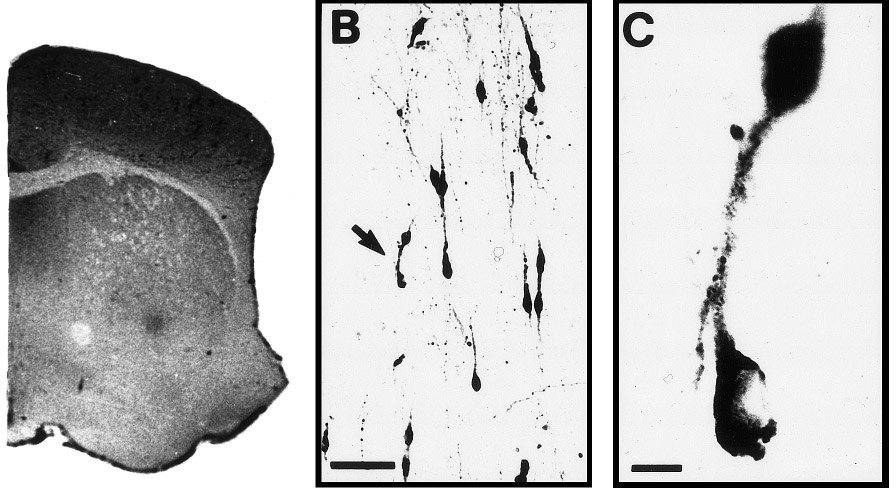
 Characteristics of the mouse GnRH forebrain system. (A) Cross-section of the mouse brain. CC, corpus callosum; AC, anterior
commissure; ON, optic nerve. Bar, 700 µm. The box indicates the area of B (not to scale). (B) A photomicrograph of representative GnRH-irneurons in the preoptic area of the mouse brain at 2 months of age. Bar, 100 µm. Arrow, GnRH-ir neurons shown in C. (C) Higher magnificationphotomicrograph of representative GnRH-ir neurons. Bar, 10 µm.
Characteristics of the mouse GnRH forebrain system. (A) Cross-section of the mouse brain. CC, corpus callosum; AC, anterior
commissure; ON, optic nerve. Bar, 700 µm. The box indicates the area of B (not to scale). (B) A photomicrograph of representative GnRH-irneurons in the preoptic area of the mouse brain at 2 months of age. Bar, 100 µm. Arrow, GnRH-ir neurons shown in C. (C) Higher magnificationphotomicrograph of representative GnRH-ir neurons. Bar, 10 µm.