He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Doi:10.1016/s1567-1348(03)00069-8
Infection, Genetics and Evolution xxx (2003) xxx–xxx
Identification of Trypanosoma brucei circulating in a sleeping
sickness focus in Cˆote d’Ivoire: assessment of genotype
Vincent Jamonneau , Christian Barnabé , Mathurin Koffi ,
Bocar Sané , Gérard Cuny , Philippe Solano
a Institut de Recherche pour le Développement (IRD), UR 035, Laboratoire de Recherche et de Coordination sur les Trypanosomoses, Programme SantéAnimale, TA 207/G, Campus Internationnal de Baillarguet, 34398 Montpellier Cedex 5, France
b Institut de Recherche pour le Développement (IRD), Génétique des Maladies Infectieuses, UR 062, UMR CNRS-IRD 9926,B.P. 64501, 34394 Montpellier Cedex 5, France
c Institut de Recherche pour le Développement (IRD), UR 035, Centre Pierre Richet, 01 B.P. 1500 Bouaké, Cˆote d’Ivoire, France
Received 27 February 2003; received in revised form 11 April 2003; accepted 11 April 2003
Abstract
Genetic studies of Trypanosoma brucei have been mainly based on rodent inoculation (RI) for isolation of trypanosome strains. However,
Trypanosoma brucei gambiense is difficult to grow in rodents. The development and use of the Kit for In Vitro Isolation (KIVI) oftrypanosomes has led to a better isolation success. However, some authors report a genetic monomorphism in T. b. gambiense, and theextensive use of the KIVI was suspected as being responsible for this low genetic diversity. In the present work, trypanosome stocks wereisolated from both humans and pigs in an active sleeping sickness focus in Cˆote d’Ivoire. Two methods were simultaneously used forthis purpose: KIVI and rodent inoculation. None of the human stocks grew in rodents. Some of the stocks originating from pigs could beisolated with both methods. Each of these stocks (from the same pig) showed a different isoenzymatic pattern according to the isolationmethod used. All the human stocks identified belonged to the major zymodeme 3 of T. b. gambiense group 1, whereas the stocks isolatedfrom pigs belonged to a new group of zymodemes even if they were genetically closely related. These observations may have significantimplications when analysing the population structure of T. brucei, and also raise again the question of the importance of the animal reservoirin Human African Trypanosomiasis (HAT). 2003 Elsevier Science B.V. All rights reserved. Keywords: Trypanosoma brucei; Isoenzyme electrophoresis; Kit for In Vitro Isolation (KIVI); Rodent inoculation; Animal reservoir; Cˆote d’Ivoire
1. Introduction
phenomenon in natural populations remains a controversialissue that needs to be clarified since it has great implications
The species Trypanosoma brucei sensu lato is classi-
on the epidemiology and control of the disease
cally divided into three subspecies on the basis of human
infectivity and geographical distribution, as they are mor-
In addition, within T. b. gambiense, acute cases have also
phologically indistinguishable: Trypanosoma brucei gambi-ense is responsible for the chronic form of Human African
to controversy on the subspecies classification of T. brucei
Trypanosomiasis (HAT) in West and Central Africa, T. b.s.l. Biochemical and molecular markers have been used ex-
rhodesiense the agent of acute form in East Africa, and T.
tensively in an attempt to establish a reliable classification. b. brucei considered as not pathogenic for humans. T. bru-
The only group which can be considered as a taxonomic
cei s.l. has been shown to undergo sexual recombination in
unit is the one defined as T. b. gambiense group 1 (
the laboratory (but the frequency of this
which represents 80% of all West and Central Africanhuman isolates. Other trypanosome strains isolated in Westand Central Africa which did not belong to this group were
∗ Corresponding author. Tel.: +33-4-67-59-39-19;
classified in a very heterogeneous T. b. gambiense group 2,
E-mail address: [email protected] (V. Jamonneau).
isolated from domestic and wild animals in these regions are
2 Tel.: +22-5-31-63-95-43; fax: +22-5-31-63-27-38.
1567-1348/03/$ – see front matter 2003 Elsevier Science B.V. All rights reserved. doi:10.1016/S1567-1348(03)00069-8
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
continued up to a maximum of 8 weeks after inoculation.
of the importance of the animal reservoir.
The positive KIVI were sub-inoculated in Cunningham’s
In most of the genetic studies of trypanosomes isolated
from human and animal mentioned earlier, isolation of try-
serum and antibiotics, according to the protocol described
panosomes has been achieved by inoculation of infected
blood to laboratory rodents and multiplication by in vivo
culture. However, due to low virulence of T. b. gambiense
For each patient, one heparinized tube of 5 ml of blood
in rodents, there has been insufficient field sampling for
was taken by venepuncture. For each pig, one heparinized
tube of 5 ml of blood was taken from the jugular vein. From
been developed for in vitro isolation of trypanosomes in the
each heparinized tube (human and pig), 0.5 ml of blood were
injected intraperitoneally in two mice. The mice were in-
medium allowed the transformation from bloodstream form
oculated with cyclophosphamide (Endoxan, 300 mg/kg) 2
to procyclic form, the later being easily cultivated on axenic
days after blood injection and then every 5 days, and were
followed up during 2 months by microscope examination
successful genetic identification of trypanosomes from hu-
of a fresh drop of blood from the tail, every 3 days from
mans, domestic and wild animals from field studies (
the 6th day after inoculation. When parasitaemia became
high (108 trypanosomes/ml), blood from the mouse was pas-
saged in rats. When the parasitaemia in the rat reached 108
biense displayed very little genetic diversity using several
trypanosomes/ml, a cardiac puncture was made, and try-
biochemical and molecular markers on a several years sam-
panosomes from the blood were filtered by using a macro
pling of trypanosomes isolated from humans in various foci
of Cˆote d’Ivoire. They suggested that the exhaustive use
of parasites were conserved into liquid nitrogen until used.
of the KIVI might be responsible for this apparent low ge-netic diversity. For instance, there has not been any stock of
2.3. Multilocus enzyme electrophoresis
the Bouafle group isolated from humans since the KIVI hasbeen used as the isolation technique. Hence, the aim of the
Proteins were extracted from the pellets according to
present work was to compare genetically by Multilocus en-
zyme electrophoresis (MLEE), trypanosomes isolated from
lose acetate plates (Helena®) and 10 enzymatic systems rep-
humans and pigs in a HAT focus in Cˆote d’Ivoire, by using
resenting 13 loci were revealed: ALAT (EC. 2.6.1.2), GOT
two isolation techniques for each sample: KIVI and rodent
(EC. 2.6.1.1), Nhi (EC. 3.2.2.1), Nhd (EC. 3.2.2.1), ME (EC.
1.1.1.40), PEP-2 (EC. 3.4.11), MDH (EC. 1.1.1.37), IDH(1.1.1.42), PGM (EC. 2.7.5.1) according to and SOD (EC. 1.15.1.1) ac-
2. Materials and methods
cording to chemicals were obtainedfrom Sigma®. For analysis procedure, UPGMA dendrogram
was built, starting with the Jaccard’s genetic distances (d,calculated from MLEE results for visualiz-
The study was undertaken in the focus of Bonon (6◦55
ing the relationships between the stocks
N, 6◦ W), in the pre-forest zone of Central-western Cˆote
Reference stocks of T. b. brucei and T. b. gambiense
d’Ivoire. From a total of 74 patients diagnosed with HAT
groups 1 and Bouafle were included, as well as stocks of
during a medical survey conducted in April 2000, the first
Trypanosoma congolense-like groups for the UPGMA com-
16 patients who gave their consent after explanation of the
parison (Reference stocks had been isolated using
objective and rationale of the study were used in this work.
the KIVI method during or after 1991, those before 1991
Patients were treated by medical staff of the National Control
Program following the protocols approved by the Ministryde la Santé of Cˆote d’Ivoire. Stocks were isolated from these16 patients using both the KIVI and RI methods (see below). 3. Results
Within the same focus, in the localities where patients werefound, blood was taken from a total of 18 pigs, and was
Of the 16 KIVI performed on human samples, 12 (pa-
inoculated in both KIVI and rodent for each of them.
tients, 1–12) gave a positive result (75%; Unfor-tunately, RI was negative for all 16 patients. Thus, stocks
of patients 13–16 could not be isolated by either KIVI orRI. Both the KIVI and RI gave a positive result for 8 out of
In the field, KIVI were kept in the shade at ambient tem-
18 pig samples (samples, 17–24). Of the 10 remaining pigs,
perature (25–30 ◦C) until arrival in the laboratory, where they
five stocks were isolated only by KIVI (samples, 25–29),
were maintained at 27 ◦C. Three examinations a week were
while five pigs gave a negative result with both KIVI and RI
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Table 1Reference stocks used for MLEE characterization with corresponding zymodeme and taxonomic identification
Kob, Kobus kob; Hartebeest, Alcelaphus buselaphus; CI, Cˆote d’Ivoire; Zym., Zymodeme; T. b. g 1, Trypanosoma brucei gambiense group 1; T. b. bfl,Trypanosoma brucei Bouafle group, T. cong., Trypanosoma congolense; Ref.: 1, 2, 3, 4,5,
(samples, 30–34). Thus, KIVI and RI were positive for 13
isolated from human by KIVI were 1-2-4, from pig by KIVI
and 8 out of 18 pigs, respectively, (72.2 and 44.4%, respec-
were 1-3-5, and from pig by RI were 1-5. The SOD system
was the most polymorphic, but the two different patterns ob-
As RI was negative for all human subjects, stocks isolated
served for SODa and the four patterns observed for SODb
by KIVI from humans were named stocks 1–12 (
were not specific for neither the host of origin nor the iso-
Stocks isolated by KIVI from pig were named “no. of the
pig KIVI” and stocks isolated by RI were named “no. of the
All the stocks isolated from humans belonged to zy-
pig Rod.”. For instance, pig no. 19, positive by both KIVI
modeme 3 (Z3), like the Sique reference stock
and RI, gave two stocks: 19 KIVI and 19 Rod.
Stocks isolated from pig by either KIVI or RI
Isoenzyme patterns of all stocks isolated are detailed in
showed different patterns corresponding to six new zy-
Stocks were grouped by zymodeme. For exam-
modemes, which we named, Z44–Z49. Within the group of
ple, 19 KIVI, 23 KIVI and 27 KIVI belonged to the same
stocks isolated from pigs with RI, nearly all were monomor-
zymodeme, Z44. Of the 13 loci analyzed, 7 (IDH, MDH,
phic except 19 Rod., which differed from the others by
ME1, NHi1, Nhi2, NHd and PGM) were monomorphic for
SODb (3-5-7 no. 3-7). Within the group of stocks isolated
all stocks. The phenotypes ALAT (6-7-8), ME2 (1-2-3) and
from pigs with KIVI, some variation occurred at loci SODa
PEP-2 (1-2-4) were specific to stocks isolated from humans.
and SODb. Each pig sample that could be isolated using the
Locus GOT gave a diagnostic pattern for each isolation
two methods (KIVI and RI) resulted in two different zy-
method (5-7-8 for KIVI and 8 for RI). Locus PEP-2 was di-
modemes according to the isolation method; this constitutes
agnostic for both isolation method and host of origin: stocks
the most important result of this study.
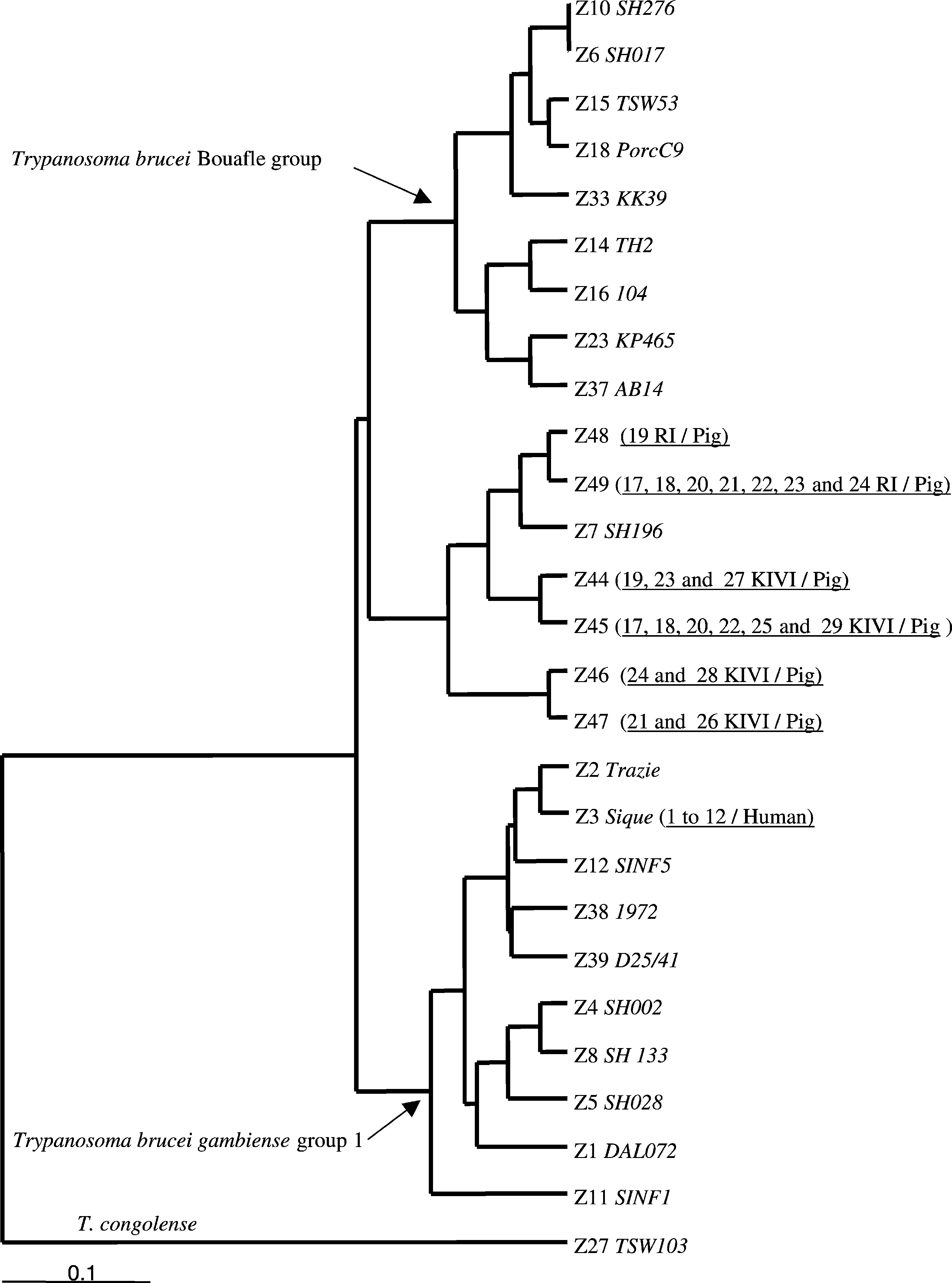
Phenetic relationships between zymodemes (including the
reference stocks and the new zymodemes) are shown in the
dendrogram (Three groups of zymodemes can be
Isolation results for both the KIVI and the rodent inoculation, performed
seen: the first one groups the Bouafle group reference stocks
on the 16 patients and the 18 pigs of our study
except Z7, the second one groups the gambiense group 1 ref-
erence stocks including Z3, and the third one is constituted
by Z7 and the new zymodemes isolated from pig using both
KIVI and RI. These new zymodemes are genetically close
to each other and seem to be different from both the bouafle
(except Z7) and the gambiense group 1 stocks. It should be
noted that this observation does not support those of
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Table 3Isoenzyme patterns of the stock isolated and corresponding zymodemes (Z.)
Rd., Rodent; Meth., Isolation method; numbers indicate presence of patterns at a given locus.
patterns were specific: (i) SODa 3-6-7-12-14, SODb 3-5
noted that the following loci were monomorphic: IDH, NHd,
for gambiense group 1, and (ii) SODa 1-2-5-11-13, SODb
NHi, PGM. We obtained the same results as theirs for the
3-7 for Bouafle group. Some stocks of our study isolated
from pig presented these two kinds of pattern (Z46 is SODa
differentiated procyclic and bloodstream forms. But, on
3-6-7-12-14, SODb 3-5 and both Z44 and Z49 are SODa
other loci we obtained different results:
1-2-5-11-13, SODb 3-7, see while they cluster out
found some polymorphism on ALAT and MDH that they still
of these two groups. It should be noted that the bootstrap
attributed to metabolic differences. In our work, MDH and
values corresponding to the three groups of zymodemes are
ALAT showed the same pattern whatever the isolation tech-
nique used, and ALAT was polymorphic but regardless of thelifecycle stage. Furthermore, we observed some polymor-phism on other loci that were not polymorphic in the work
4. Discussion
of ME, PEP-2, SOD. Since differentpatterns were also observed on stocks isolated by the same
In this work carried out in Cˆote d’Ivoire, stocks from
technique, we may conclude that this polymorphism was
sleeping sickness patients and pigs from the same area were
not only due to differences of trypanosome lifecycle stages
isolated simultaneously by two isolation methods, KIVI
but probably corresponds to an intrinsic genetic variation.
and RI, in order to study a possible selective effect of the
Our hypothesis is that these pigs were probably infected
use of KIVI, which had been suspected in previous works
by two or more different genotypes of T. brucei s.l. which
were selected by the isolation method. It might be possi-
from pigs could be isolated by the two methods and a differ-
ble that the same phenomenon has occurred in the work of
ence of banding pattern was observed between stocks iso-
lated by KIVI and stocks isolated by RI from the same pig.
ported in experimental infections of rodents: when two dif-
Here, the two isolation methods used implicate different
ferent stocks are inoculated in a mouse, one of them grows
lifecycle stages (procyclic forms for KIVI, bloodstream for
at the expense of the other (Further investiga-
RI). It should be noted that other authors found differences
tions are required in order to confirm this result, especially,
in isoenzyme patterns within T. b. gambiense which they at-
tributed to metabolic differences between trypanosome life-
Intra-specific multiple infections of T. brucei s.l. have al-
if the differences observed in the present work could not be
due only to physiological differences between procyclic and
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Fig. 1. The reference stocks are in italics and are preceded by their zymodeme number. The stocks obtained in the present study are underlined, are alsopreceded by their zymodeme number, and are followed by the host of origin (e.g. sample Z48 (19 RI/pig), is sample number 19, isolated from pig byrodent inoculation, and belongs to zymodeme 48).
Mixed infections of two genotypes of T. b.
by this method, justifying the suspicion of a selective effect
gambiense group 1 have been recently observed by
man stocks. Unfortunately, this study did not help us to an-
tion of stocks by KIVI from the same patient at two dif-
swer this question because none of the human stocks grew
ferent times (these two stocks were identified as zymodeme
in rodents, thus confirming the usually low isolation success
3 by MLEE). Indeed, in this latter work, MLEE identifi-
of RI for T. b. gambiense, which justified the use of KIVI.
cation of the stocks isolated from human with KIVI con-
The selective effect of the isolation and/or cultivation tech-
firmed the unusual genetic homogeneity of stocks isolated
niques has already been reported in bacteria and also in other
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
parasitic diseases, e.g. Chagas disease (Trypanosoma cruzy)
supported by the Institut de Recherche pour le Développe-
ment (IRD), by the Agence Française de la Francophonie
maniasis (Leishmania donovani; In HAT,
(AUF), and by the French Ministère de l’Education Na-
this selective effect had been already suspected in several
tionale et de la Recherche. We thank G. Manners for
studies (e.g. but has not been really in-
The selective effect of isolation techniques has important
References
implications in term of population genetics of the organismstudied and on the epidemiology of the corresponding dis-
Aerts, D., Truc, P., Penchenier, L., Claes, Y., Le Ray, D., 1992. A Kit for
ease. For example, in HAT, if the trypanosomes stocks under
In Vitro Isolation of trypanosomes in the field: first trial with sleeping
study were isolated by using only one method, a clonal struc-
sickness patients in the Congo. Trans. R. Soc. Trop. Med. Hyg. 86,
ture with only one genotype might arise (e.g. zymodeme 3
of T. b. gambiense with KIVI in West Africa,
Bosseno, M.F., Yacsik, N., Vargas, F., Breniere, S.F., 2000. Selection of
whereas the real population structure remains
Trypanosoma cruzy clonal genotypes (clonet 20 and 39) isolated fromBolivian tryatomines following subculture in liquid medium. Mem.
unknown. This has been termed ‘iceberg bias’ (
This problem of sampling might be resolved by trying
Breniere, S.F., Bosseno, M.F., Revollo, S., Rivera, M.T., Carlier, Y.,
to directly identify trypanosomes from the biological fluids
Tibayrenc, M., 1992. Direct identification of Trypanosoma cruzy natural
of humans (blood, CSF and lymph fluid) without isolation
clones in vectors and mammalian hosts by polymerase chain reaction
and culture, and by using specific molecular markers. Re-
amplification. Am. J. Trop. Med. Hyg. 46, 335–341.
Cunningham, I., 1977. New culture medium for maintenance of tsetse
cently, developed markers, such as micro- or mini-satellites
tissues and growth of trypanosomatids. J. Protozool. 21, 325–329.
Dey, T., Afrin, F., Anam, K., Ali, N., 2002. Infectivity and virulence
of Leishmania donovani promastigotes: a role for media, source, and
The stocks isolated from pigs with both KIVI and RI
strain of parasite. J. Eukaryot. Microbiol. 49, 270–274.
represented six new zymodemes, which clustered together.
Dukes, P., McNamara, J.J., Godfrey, D.G., 1991. Elusive trypanosomes.
Ann. Trop. Med. Parasitol. 85, 21–32.
Therefore, whatever the isolation method, stocks isolated
Gashumba, J.K., Baker, R.D., Godfrey, D.G., 1988. Trypanosoma
from pigs were different from the stocks isolated from hu-
congolense: the distribution of enzymatic variants in East and West
mans. Hence, from the results of this work, pigs might
not be incriminated as an animal reservoir of trypanosomes
Gibson, W.C., 1986. Will the real Trypanosoma brucei gambiense please
pathogenic to humans. Conversely, in Cameroon,
stand up. Parasitol. Today 2, 255–257.
Godfrey, D.G., Baker, R.D., Rickman, L.R., Mehlitz, D., 1990. The
distribution, relationships and identification of enzymatic variants
to humans was also found in pigs. However, in our study,
within the subgenus Trypanozoon. Adv. Parasitol. 29, 1–39.
zymodeme Z7 isolated from humans, which groups in the
Jaccard, P., 1908. Nouvelles recherches sur la distribution florale. Bull.
dendrogram with the new group of zymodemes isolated
Soc. Vaudoise Sci. Nat. 44, 223–270.
Jamonneau, V., N’Guessan, P., N’Dri, L., Simaro, P., Truc, P., 2000.
from pigs, had previously been classified as Bouafle group
Exploration of the distribution of Trypanosoma brucei ssp. in West
Africa, by multilocus enzyme electrophoresis. Ann. Trop. Med.
responding to the three groups of zymodemes here are not
significant. This means that the stocks isolated from pigs
Jamonneau, V., Garcia, A., Ravel, S., Cuny, G., Oury, B., Solano, P.,
are genetically very close to the ‘human’ stocks. In addi-
N’Guessan, P., N’Dri, L., Sanon, R., Frézil, J.L., Truc, P., 2002. Geneticcharacterization of Trypanosoma brucei ssp. and clinical evolution of
tion, while the group of stocks from pigs does not seem to
Human African Trypanosomiasis in Cˆote d’Ivoire. Trop. Med. Int.
belong neither to T. b. gambiense group 1 nor to the Bouafle
group in the dendrogram, some of them presented SODa
Jenni, L., Marti, J., Schweizer, J., Betschart, B., Le Page, R.W.F., Wells,
and SODb patterns previously described as specific to these
J.M., Tait, A., Paindavoine, P., Pays, E., Steinert, M., 1986. Hybrid
formation between African trypanosomes during cyclical transmission. Nature 322, 173–175.
as the blood infectivity incubation test (BIIT,
Kaukas, A., Gashumba, J.K., Lanham, S.M., Dukes, P., 1990. The
was not performed, and considering the hy-
substitution of procyclic for bloodstream form Trypanosoma brucei
pothesis of a selective effect of the KIVI and the limited
gambiense in isoenzyme studies. Trans. R. Soc. Trop. Med. Hyg. 84,
number of pigs sampled, it is difficult to conclude the role of
pigs in terms of animal reservoir in our study area. Further
Lumsden, W.H.R., Kimber, C.D., Strange, M., 1977. Trypanosoma brucei:
a miniature anion exchange/centrifugation technique for the detection
studies are in progress to shed light on this issue.
of low parasitemias in mice. Trans. R. Soc. Trop. Med. Hyg. 71, 421–424.
Macleod, A., Turner, C.M.R., Tait, A., 1999. A high level of mixed
Acknowledgements Trypanosoma brucei infections in tsetse flies detected by threehypervariable mini-satellites. Mol. Biochem. Parasitol. 102, 237–248.
Macleod, A., Tweedie, A., Welburn, S.C., Maudlin, I., Turner, C.M.R.,
We thank the HAT team of the Institut Pierre Richet in
Tait, A., 2000. Mini-satellite marker analysis of Trypanosoma brucei:
Bouaké and the National Control Program for HAT of Cˆote
reconciliation of clonal, panmictic, and epidemic population genetic
d’Ivoire for their excellent technical support. This work was
structures. Proc. Natl. Acad. Sci. U.S.A. 97, 13442–13447. V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxxLeishmania, Naegleria, Plasmodium, Trichomonas and Trypanosoma
Epidemiological studies on the animal reservoir of gambiense
and their medical and taxonomical consequences. Proc. Natl. Acad.
sleeping sickness. Part III. Characterization of Trypanozoon stocks by
isoenzymes and sensitivity to human serum. Tropenmed. Parasitol. 33,
trypanosomiase humaine Africaine: intérˆet de l’identification génétique
Nkinin, S.W., Njiokou, F., Penchenier, L., Grebaut, P., Simo, G., Herder,
des trypanosomes. Med. Trop. 61, 433–436.
S., 2002. Characterization of Trypanosoma brucei s. l. subspecies by
Truc, P., Tibayrenc, M., 1993. Population genetics of Trypanosoma
isoenzymes in domestic pigs from the Fontem sleeping sickness focus
brucei in Central Africa: taxonomic and epidemiological significance.
of Cameroon. Acta Trop. 81, 225–232.
Rickman, L.R., Robson, J., 1970. The testing of proven Trypanosoma
Truc, P., Mathieu-Daude, F., Tibayrenc, M., 1991. Multilocus isoenzyme
brucei and T. rhodesiense strains by the blood incubation infectivity
identification of Trypanosoma brucei stocks isolated in Central Africa:
test. Bull. World Health Organ. 42, 911–916.
evidence for an animal reservoir of sleeping sickness in Congo. Acta
Scott, C.M., 1981. Mixed populations of Trypanosoma brucei in a naturally
infected pig. Tropenmed. Parasitol. 32, 221–222.
Truc, P., Aerts, D., Mcnamara, J.J., Claes, Y., Allimgham, R., Le Ray,
Seed, J.R., 1978. Competition among serologically different clones of
D., Godfrey, D.G., 1992. The direct in vitro isolation of TrypanosomaTrypanosoma brucei gambiense in vivo. J. Protozool. 25, 526–529. brucei from man and animals, and its potential value for the diagnosis
Sneath, P.H.A., Sokal, R.R., 1973. Numerical Taxonomy. The Principles
of Gambian trypanosomiasis. Trans. R. Soc. Trop. Med. Hyg. 86, 627–
and Practice of Numerical Classification. In: Kennedy, D., Park, R.B.
Truc, P., Formenty, P., Diallo, P.B., Komoin-Oka, C., Lauginie, F., 1997a.
Stevens, J.R., Nunes, V.L.B., Lanham, S.M., Oshiro, E.T., 1989.
Confirmation of two distinct classes of zymodemes of Trypanosoma
Isoenzyme characterization of Trypanosoma evansi isolated from
brucei infecting patients and wild mammals in Cˆote d’Ivoire: suspected
capybaras and dogs in Brazil. Acta Trop. 46, 213–222.
difference in pathogenicity. Ann. Trop. Med. Parasitol. 91, 951–956.
Stevens, J.R., Lanham, S.M., Allingham, R., Gashumba, J.K., 1992. A
Truc, P., Formenty, P., Duvallet, G., Komoin-Oka, C., Diallo, P.B.,
simplified method for identifying subspecies and strain groups in
Lauginie, F., 1997b. Identification of trypanosomes isolated by KIVI
Trypanozoon by isoenzymes. Ann. Trop. Med. Parasitol. 86, 9–28.
from wild mammals in Cˆote d’Ivoire: diagnostic, taxonomic and
Stevens, J.R., Mathieu-Daude, F., Mcnamara, J.J., Mizen, V.H., Nzila,
epidemiological considerations. Acta Trop. 67, 187–196.
A., 1994. Mixed populations of Trypanosoma brucei in wild Glossina
Truc, P., Ravel, S., Jamonneau, V., N’Guessan, P., Cuny, G., 2002. Genetic
palpalis palpalis. Trop. Med. Parasitol. 45, 313–318.
variability within Trypanosoma brucei gambiense: evidence for the
Tibayrenc, M., 1999. Toward an integrated genetic epidemiology of
circulation of different genotypes in Human African Trypanosomiasis
parasitic protozoa and other pathogens. Ann. Rev. Genet. 33, 449–
patients in Cˆote d’Ivoire. Trans. R. Soc. Trop. Med. Hyg. 96, 1–4.
Young, C.J., Godfrey, D.G., 1983. Enzyme polymorphism and the
Tibayrenc, M., Kjellberg, F., Ayala, F.J., 1990. A clonal theory of
distribution of T. congolense isolates. Ann. Trop. Med. Parasitol. 77,
parasitic protozoa: the population structures of Entamoeba, Giardia,
EYE INJURIES IN BOXING Jean-Louis Llouquet Introduction The era of blindness as a result of boxing is past. However boxing doctors have become more aware of ocular damage: retinal detachment being the most frequently observed serious injury in professional boxing. Ocular injuries as a result of boxing mainly affects professional boxers The eye is relatively well pr
Nr. 6, oktober 1997 Nieuwsbrief neuromusculaire ziekten V ereniging AMYOTROFISCHE Amyotrofische lateraal sclerose (ALS) is een progressieve aandoening die leidt tot krachtsverlies LATERAAL van de extremiteiten, romp- en ademhalingsmusculatuur en van de slik-, kauw- en SCLEROSE spraakspieren. Naast medicamenteuze behandeling met riluzole is de behandeling van ALS voorname



 Infection, Genetics and Evolution xxx (2003) xxx–xxx
Identification of Trypanosoma brucei circulating in a sleeping
sickness focus in Cˆote d’Ivoire: assessment of genotype
Vincent Jamonneau , Christian Barnabé , Mathurin Koffi ,
Bocar Sané , Gérard Cuny , Philippe Solano
a Institut de Recherche pour le Développement (IRD), UR 035, Laboratoire de Recherche et de Coordination sur les Trypanosomoses, Programme Santé
Animale, TA 207/G, Campus Internationnal de Baillarguet, 34398 Montpellier Cedex 5, France
b Institut de Recherche pour le Développement (IRD), Génétique des Maladies Infectieuses, UR 062, UMR CNRS-IRD 9926,
B.P. 64501, 34394 Montpellier Cedex 5, France
c Institut de Recherche pour le Développement (IRD), UR 035, Centre Pierre Richet, 01 B.P. 1500 Bouaké, Cˆote d’Ivoire, France
Received 27 February 2003; received in revised form 11 April 2003; accepted 11 April 2003
Abstract
Infection, Genetics and Evolution xxx (2003) xxx–xxx
Identification of Trypanosoma brucei circulating in a sleeping
sickness focus in Cˆote d’Ivoire: assessment of genotype
Vincent Jamonneau , Christian Barnabé , Mathurin Koffi ,
Bocar Sané , Gérard Cuny , Philippe Solano
a Institut de Recherche pour le Développement (IRD), UR 035, Laboratoire de Recherche et de Coordination sur les Trypanosomoses, Programme Santé
Animale, TA 207/G, Campus Internationnal de Baillarguet, 34398 Montpellier Cedex 5, France
b Institut de Recherche pour le Développement (IRD), Génétique des Maladies Infectieuses, UR 062, UMR CNRS-IRD 9926,
B.P. 64501, 34394 Montpellier Cedex 5, France
c Institut de Recherche pour le Développement (IRD), UR 035, Centre Pierre Richet, 01 B.P. 1500 Bouaké, Cˆote d’Ivoire, France
Received 27 February 2003; received in revised form 11 April 2003; accepted 11 April 2003
Abstract V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
continued up to a maximum of 8 weeks after inoculation.
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
continued up to a maximum of 8 weeks after inoculation. V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Table 1Reference stocks used for MLEE characterization with corresponding zymodeme and taxonomic identification
Kob, Kobus kob; Hartebeest, Alcelaphus buselaphus; CI, Cˆote d’Ivoire; Zym., Zymodeme; T. b. g 1, Trypanosoma brucei gambiense group 1; T. b. bfl,Trypanosoma brucei Bouafle group, T. cong., Trypanosoma congolense; Ref.: 1, 2, 3, 4,5,
(samples, 30–34). Thus, KIVI and RI were positive for 13
isolated from human by KIVI were 1-2-4, from pig by KIVI
and 8 out of 18 pigs, respectively, (72.2 and 44.4%, respec-
were 1-3-5, and from pig by RI were 1-5. The SOD system
was the most polymorphic, but the two different patterns ob-
As RI was negative for all human subjects, stocks isolated
served for SODa and the four patterns observed for SODb
by KIVI from humans were named stocks 1–12 (
were not specific for neither the host of origin nor the iso-
Stocks isolated by KIVI from pig were named “no. of the
pig KIVI” and stocks isolated by RI were named “no. of the
All the stocks isolated from humans belonged to zy-
pig Rod.”. For instance, pig no. 19, positive by both KIVI
modeme 3 (Z3), like the Sique reference stock
and RI, gave two stocks: 19 KIVI and 19 Rod.
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Table 1Reference stocks used for MLEE characterization with corresponding zymodeme and taxonomic identification
Kob, Kobus kob; Hartebeest, Alcelaphus buselaphus; CI, Cˆote d’Ivoire; Zym., Zymodeme; T. b. g 1, Trypanosoma brucei gambiense group 1; T. b. bfl,Trypanosoma brucei Bouafle group, T. cong., Trypanosoma congolense; Ref.: 1, 2, 3, 4,5,
(samples, 30–34). Thus, KIVI and RI were positive for 13
isolated from human by KIVI were 1-2-4, from pig by KIVI
and 8 out of 18 pigs, respectively, (72.2 and 44.4%, respec-
were 1-3-5, and from pig by RI were 1-5. The SOD system
was the most polymorphic, but the two different patterns ob-
As RI was negative for all human subjects, stocks isolated
served for SODa and the four patterns observed for SODb
by KIVI from humans were named stocks 1–12 (
were not specific for neither the host of origin nor the iso-
Stocks isolated by KIVI from pig were named “no. of the
pig KIVI” and stocks isolated by RI were named “no. of the
All the stocks isolated from humans belonged to zy-
pig Rod.”. For instance, pig no. 19, positive by both KIVI
modeme 3 (Z3), like the Sique reference stock
and RI, gave two stocks: 19 KIVI and 19 Rod. V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Table 3Isoenzyme patterns of the stock isolated and corresponding zymodemes (Z.)
Rd., Rodent; Meth., Isolation method; numbers indicate presence of patterns at a given locus.
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Table 3Isoenzyme patterns of the stock isolated and corresponding zymodemes (Z.)
Rd., Rodent; Meth., Isolation method; numbers indicate presence of patterns at a given locus.
 V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Fig. 1. The reference stocks are in italics and are preceded by their zymodeme number. The stocks obtained in the present study are underlined, are alsopreceded by their zymodeme number, and are followed by the host of origin (e.g. sample Z48 (19 RI/pig), is sample number 19, isolated from pig byrodent inoculation, and belongs to zymodeme 48).
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Fig. 1. The reference stocks are in italics and are preceded by their zymodeme number. The stocks obtained in the present study are underlined, are alsopreceded by their zymodeme number, and are followed by the host of origin (e.g. sample Z48 (19 RI/pig), is sample number 19, isolated from pig byrodent inoculation, and belongs to zymodeme 48). V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
parasitic diseases, e.g. Chagas disease (Trypanosoma cruzy)
supported by the Institut de Recherche pour le Développe-
ment (IRD), by the Agence Française de la Francophonie
maniasis (Leishmania donovani; In HAT,
(AUF), and by the French Ministère de l’Education Na-
this selective effect had been already suspected in several
tionale et de la Recherche. We thank G. Manners for
studies (e.g. but has not been really in-
The selective effect of isolation techniques has important
References
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
parasitic diseases, e.g. Chagas disease (Trypanosoma cruzy)
supported by the Institut de Recherche pour le Développe-
ment (IRD), by the Agence Française de la Francophonie
maniasis (Leishmania donovani; In HAT,
(AUF), and by the French Ministère de l’Education Na-
this selective effect had been already suspected in several
tionale et de la Recherche. We thank G. Manners for
studies (e.g. but has not been really in-
The selective effect of isolation techniques has important
References V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Leishmania, Naegleria, Plasmodium, Trichomonas and Trypanosoma
Epidemiological studies on the animal reservoir of gambiense
and their medical and taxonomical consequences. Proc. Natl. Acad.
V. Jamonneau et al. / Infection, Genetics and Evolution xxx (2003) xxx–xxx
Leishmania, Naegleria, Plasmodium, Trichomonas and Trypanosoma
Epidemiological studies on the animal reservoir of gambiense
and their medical and taxonomical consequences. Proc. Natl. Acad.