He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Ruifreitas.info
Diet of Breeding Ospreys in the Cape Verde Archipelago, Northwestern Africa
Author(s) :Samir Martins, Rui Freitas, Luís Palma, and Pedro BejaSource: Journal of Raptor Research, 45(3):244-251. 2011. Published By: The Raptor Research FoundationDOI: 10.3356/JRR-10-101.1
BioOne () is a a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainableonline platform for over 170 journals and books published by nonprofit societies,associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should bedirected to the individual publisher as copyright holder.
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
J. Raptor Res. 45(3):244–251E 2011 The Raptor Research Foundation, Inc.
DIET OF BREEDING OSPREYS IN THE CAPE VERDE ARCHIPELAGO,
Departamento de Engenharias e Cieˆncias do Mar, Universidade de Cabo Verde, CP 163 Mindelo,
CIBIO, Centro de Investigac¸a˜o em Biodiversidade e Recursos Gene´ticos, Campus Agra´rio de Vaira˜o,
Universidade do Porto, Vaira˜o 4485-601, Portugal
ABSTRACT.—We studied the diet of breeding Ospreys (Pandion haliaetus) in the Cape Verde archipelagoduring 2006, using prey remains recovered at 21 nests and perches on the islands of Sa˜o Vicente, Santiago,Santa Luzia, Boavista, Branco, and Raso. We identified a total of 1264 individual fish prey items of 35species. Diet was dominated (.80%) by only six fish species, including Trachinotus ovatus, Exocoetus volitans,Aulostomus strigosus, Sparisoma cretense, Sardinella maderensis, and Tylosurus acus. Dominant prey species variedamong islands, but diet similarity was greater between nearby islands. Pelagic species were consumed mostfrequently (.60%) in Boavista and Santa Luzia, whereas demersal reef fish dominated (.50%) in theother islands. The fish consumed were generally large, though there was wide variation in estimated length(20.7–62.2 cm) and weight (49–1117 g). A comparison of Osprey diet with Cape Verde fisheries suggestedthat the potential for conflict is low, due to minimal overlap in the primary species caught. Changes inmarine productivity associated with the ongoing moderate warming of the Canary Current System mayrepresent a threat, though there is considerable uncertainty about the type and magnitude of these effects. Monitoring of Osprey numbers, breeding success, and diet is required to detect any changes associated withavailability of food sources, and such monitoring may also provide a relatively simple and inexpensivemethod to track long-term changes in littoral fish assemblages.
Osprey; Pandion haliaetus; Cape Verde; diet; foraging ecology; piscivory.
DIETA DE INDIVIDUOS REPRODUCTIVOS DE PANDION HALIAETUS EN EL ARCHIPIE
RESUMEN.—Estudiamos la dieta de individuos reproductivos de Pandion haliaetus en el archipie´lago de CaboVerde durante 2006, usando restos de presas recolectados en 21 nidos y perchas en las islas Sa˜o Vicente,Santiago, Santa Luzia, Boavista, Branco y Raso. Identificamos un total de 1264 presas de peces correspon-dientes a 35 especies. La dieta estuvo dominada (.80%) por so´lo seis especies de peces, incluyendoTrachinotus ovatus, Exocoetus volitans, Aulostomus strigosus, Sparisoma cretense, Sardinella maderensis y Tylosurusacus. Las especies dominantes de presas variaron entre las islas, pero la similitud de las dietas fue mayorentre las islas cercanas. Las especies pela´gicas fueron consumidas con mayor frecuencia (.60%) en Boa-vista y Santa Luzia, mientras que los peces de fondo de los arrecifes dominaron (.50%) en otras islas. Lospeces consumidos fueron generalmente grandes, aunque hubo gran variacio
peso (49–1117 g) estimado. Una comparacio
´ n de la dieta de P. haliaetus con las pesqueras de Cabo Verde
sugirio´ que existe poco potencial de conflicto, debido a la baja superposicio´n en la principal especiecapturada. Los cambios en la productividad marina asociados con el calentamiento moderado en cursodel Sistema de Corriente de las Canarias pueden representar una amenaza, aunque hay una incertidumbreconsiderable sobre el tipo y la magnitud de estos efectos. Se requiere el monitoreo de los nu
e´xito reproductivo y de la dieta de P. haliaetus para detectar cualquier cambio asociado con la disponibi-lidad de las fuentes de alimento, y este monitoreo tambie´n puede brindar un me´todo relativamente simpley barato para seguir los cambios a largo plazo en los ensambles de peces de litoral.
1 Present address: Cabo Verde Natura 2000, Rua 1u de Maio, CP 100 Sal Rei, Cabo Verde. 2 Email address: [email protected]
The Osprey (Pandion haliaetus) has a nearly world-
mentioned in later studies (Naurois 1987, Ontiveros
wide distribution and large population sizes (Poole
2003). Instead, fish of the families Carangidae,
1989), despite dramatic, but transitory, reductions
Acanthuridae, Scombridae, and Exocoetidae were
in range and numbers due to organochlorine-in-
obtained from nests in Santa Luzia and Raso (Naur-
duced reproductive suppression during the second
ois 1987), and parrotfish of the genus Sparisoma
half of the twentieth century (Watts and Paxton
were by far the most important prey recorded in
2007, Bai et al. 2009). The global conservation status
Boavista (Ontiveros 2003). These preliminary data
of Ospreys is thus of Least Concern (BirdLife Inter-
suggest that Ospreys in the Cape Verdes may exploit
national 2010), though there is evidence that some
a wide range of marine fish, though there may be
local populations are highly vulnerable to extermi-
strong spatial variation in the most important prey
nation. This is the case for small and isolated pop-
species. We here provide a comprehensive account
ulations in the Mediterranean and Macaronesia,
of Osprey diet in the Cape Verde archipelago. Spe-
where the species is restricted to the islands of Cor-
cifically, we describe: (1) prey species and sizes most
sica and the Baleares, the North African coast, and
frequently consumed by Ospreys, (2) dietary varia-
the Atlantic archipelagos of the Canaries and Cape
tion across islands, and (3) the fish habitats (pelagic
Verde (Thibault et al. 1996, Palacios 2004, Palma et
vs. demersal) most often exploited by Ospreys. We
al. 2004). Protection of peripheral populations such
use our results to identify key trophic resources for
as these may be important for the long-term conser-
Ospreys in the Cape Verde islands and to discuss the
vation of even geographically widespread species
conservation implications of potential variations in
resource availability due to fisheries and climate
With about 70–80 breeding pairs, the Cape Verde
archipelago holds the largest Osprey population inthe whole of the Mediterranean and Macaronesia
(Palma et al. 2004). Ospreys are largely concentrat-
Study Area. The Cape Verde archipelago is made
ed in the northern Barlavento (windward) islands
up of 10 islands and several islets of volcanic origin
group (Santo Anta˜o, Sa˜o Vicente, Santa Luzia, Sa˜o
(surface: 4026 km2; coastline: 1050 km) between
Nicolau, Branco, Raso, Boavista, and Sal), where the
14u509–17u209N and 22u409–25u309W, about 600 km
population appears to be recovering from past de-
west of the African mainland (Senegal). Maximum
clines due to overharvesting of eggs and nestlings
distance between islands (Santo Anta˜o–Maio) is
for human consumption (Palma et al. 2004). In the
about 280 km. All islands are inhabited except Santa
southern Sotavento (leeward) islands (Maio, San-
Luzia and the islets. Cape Verde is included in the
tiago, Fogo, Rombos, and Brava) the species is
African Sahelian arid and semiarid climate regions
scarce and seemingly still declining (Palma et al.
and in the Macaronesian biogeographic region
2004). Reasons for these contrasting population siz-
(Duarte and Romeiras 2009). The climate is dry
es and trends are poorly known, but may be related
tropical, with roughly three main seasons: warm
at least partly to differences in prey availability
dry (May–July), warm wet (August–September),
(Naurois 1987). The Barlavento islands are influ-
and cold dry (December–April; Medina et al.
enced by the Canary Current System, which brings
2007). Ospreys breed during the cold dry season
relatively cold waters rich in nutrients from upwell-
(Naurois 1987), when water surface temperature is
ing areas in northern Africa (Roy and Cury 2003).
This creates the conditions for high marine produc-
Dietary Analysis. We collected fish remains recov-
tivity and fish abundance (Roy and Cury 2003),
ered from 21 nests and perch sites in January 2006
which in turn may influence Osprey population dis-
(Branco and Raso islets) and between April and
tribution, density, and breeding success (Harmata
June 2006 (islands of Sa˜o Vicente, Santiago, Santa
et al. 2007, Bai et al. 2009, Glass and Watts 2009).
Luzia, and Boavista). Sampling was limited by the
Despite the value of foraging ecology for under-
number of nests in each island (Palma et al. 2004),
standing population dynamics and spatial distribu-
by difficulties of access of remote nests and perches,
tion patterns, little is known about Osprey diet in
and by travel constraints between and within is-
the Cape Verde islands. Early reports described the
lands. Fish remains collected at each sampling site
comber (Serranus cabrilla) as the most important
were stored in plastic bags, labeled, and frozen for
prey of Ospreys in the archipelago (Bannerman
future analysis. We identified remains using a refer-
and Bannerman 1968), but this species was not
ence collection of bones and scales of fish occurring
in the Cape Verde archipelago (e.g., Reiner 1996),
67.8%) than demersal species (32.2%; 95% CI:
obtained from local fishermen and marine biolo-
32.2–37.4%). The only non-fish items recorded
gists. We then estimated the minimum number of
were one juvenile cat (Felis catus), one Bulwer’s Pe-
individuals of each species represented in a sample
trel (Bulweria bulwerii), and one Little Shearwater
from the number and sizes of diagnostic bones
(Puffinus assimilis boydi), although it was uncertain
(e.g., maxillae, premaxillae, operculae, etc.). We al-
whether these items were taken as prey or as nest
so used bones to estimate the size of fish captured
by Ospreys, from regression equations relating bone
Comparisons Among Islands. The number of prey
size (e.g., width, breadth, maximum length) to fish
species recorded per island varied widely across the
length and length-weight regressions (S. Martins un-
archipelago (Table 2), but there was a tendency for
this to be correlated with variation in sample sizes
Data Analysis. We quantified diet as the numeric
among islands, albeit not significantly so (r 5 0.80,
frequency of each prey type, computed as the num-
n 5 5, P 5 0.105). Diet diversity was uncorrelated
ber of individuals of that prey type relative to the
with sample sizes (r 5 0.29, n 5 5, P 5 0.633), with
total number of individuals identified (e.g., Beja
the lowest value on Santiago and the highest in the
1997). To assess whether Ospreys were feeding on
northern group of islands, particularly on Santa Lu-
fish living mainly in the water column or near the
sea bottom, we classified species as either pelagic or
For further analyses of diet composition, we com-
demersal according to their main habitat type (Fro-
bined fish with percentage numbers ,5% in the
ese and Pauly 2010). We used contingency tables
category ‘‘other fish’’ (16.6% of individuals identi-
(Legendre and Legendre 1998) to assess variation
fied). We also combined data from the nearby islets
in diet composition among islands. We also used
of Branco and Raso (about 6 km), due to small
correspondence analysis (CA) to summarize the
sample sizes. Analysis revealed a significant dietary
spatial patterns of dietary variation (Legendre and
variation (x2 5 639.0, df 5 24, P , 0.001), with
Legendre 1998). Before CA, we square-root-trans-
major differences among islands in dominant prey
formed proportional diet composition data to re-
species (Table 2). The first two axes extracted from
duce the influence of a few prey species that were
correspondence analysis accounted for 79% of var-
taken very frequently (Legendre and Legendre
iation in the data, showing a strong segregation be-
1998). We estimated diet diversity using Shannon’s
tween the southern (Santiago), central (Boavista),
entropy H (Legendre and Legendre 1998).
and northern (Sa˜o Vicente, Santa Luzia, andBranco/Raso) islands (Fig. 1).
Santiago was characterized by a diet largely dom-
Overall Diet Patterns. We recovered prey remains
inated by A. strigosus, with T. ovatus and Sardinella
from Osprey nests and perches representing 1264
maderensis also making an important dietary contri-
individual fishes and 32 species of 24 families (Ta-
bution. More than half the fish consumed at Boa-
ble 1). Most remains originated from the islands of
vista were T. ovatus, though E. volitans and several
Boavista (38.3%), Sa˜o Vicente (37.0%), and Santa
minor prey species (18) also were taken frequently.
Luzia (18.4%), whereas few were gathered on San-
Diet in the northern group was mostly characterized
tiago (1.5%), Raso (4.0%), and Branco (0.7%). De-
by the high contribution of prey species that were
spite the large number of families recorded, nearly
generally less represented on Boavista and Santiago,
80% of fish prey belonged to just five families: Car-
including E. volitans, S. cretense, and T. acus. Howev-
angidae (26.8%), Exocoetidae (16.6%), Aulostomi-
er, there were important dietary differences within
dae (14.6%), Scaridae (11.2%), and Clupeidae
the northern group, with A. strigosus dominating on
(10.2%). Likewise, six species with numeric fre-
Sa˜o Vicente, S. cretense and T. acus in Santa Luzia,
quencies .5% accounted for 83.5% of fish identi-
and the ‘‘other fish’’ in Branco/Raso.
fied, including the pompano (Trachinotus ovatus),
The proportion of pelagic versus demersal species
the tropical two-wing flying fish (Exocoetus volitans),
consumed also varied among islands (x2 5 172.6, df
the Atlantic cornetfish (Aulostomus strigosus), the
5 4, P , 0.001), with pelagic fish dominating in
parrotfish (Sparisoma cretense), the Madeiran sardi-
Boavista (86.0%) and Santa Luzia (64.8%). Pelagic
nelle (Sardinella maderensis), and the agujon needle-
fish were less important than demersal fish on Sa˜o
fish (Tylosurus acus; Table 1). Pelagic fish were far
Vicente (47.9%), Santiago (42.1%), and the islets of
more frequently consumed (65.2%; 95% CI: 62.6–
Fish prey species recorded in the diet of Ospreys in the Cape Verde archipelago (January–June 2006),
indicating fish family, the main habitat type (P 5 pelagic; D 5 demersal), the percentage of islands where it occurredin the diet (% islands), and the frequency in the diet (% numbers).
Fish Sizes. We obtained data on lengths and
sumed a wide variety of pelagic and demersal fish,
weights of 162 individual fish prey, including the
but only a few species were important in the diet.
four most important species and the surgeonfish
This was particularly evident at the scale of indi-
(Acanthurus monroviae; Table 3). Fish sizes varied
vidual islands, with just 1–3 species accounting for
widely, with estimated lengths between 20.7 and
.50% of prey consumed on each island. These key
62.2 cm (mean: 38.6 6 8.6 cm [SD]), and weights
prey species varied greatly across the archipelago,
between 49 and 1117 g (mean: 366 6 211 g [SD]).
though there was a tendency for higher dietary sim-
The longest fish captured tended to be A. strigosus,
ilarity among nearby islands, probably due to differ-
but they usually had a low mass. The heaviest fish
entiation in fish assemblage structure associated
with geographic distance (Medina et al. 2007).
Most fish prey recorded at Cape Verde have been
rarely or never found in Osprey diets elsewhere,
As in other studies (e.g., Poole 1989, Francour
likely reflecting the specificity of fish communities
and Thibault 1996, Cartron and Molles 2002, Clancy
around the archipelago (Floeter et al. 2008). Nev-
2005), Ospreys in the Cape Verde archipelago con-
ertheless, the species most frequently consumed at
Variation in frequency (% of total numbers), number of prey species, and diversity of Osprey diet among
islands of the Cape Verde archipelago (January–June 2006); n 5 number of fish prey items identified.
FREQUENCY OF FISH SPECIES IN THE DIET (%) BY ISLAND
Cape Verde shared some ecological and morpho-
dant where the rocky coast plunges abruptly to the
logical similarities with those taken by Ospreys feed-
depths (Safriel et al. 1985), which may explain their
ing in other subtropical and tropical marine waters.
high consumption in islands with very narrow rocky
For example, the fish most commonly captured by
Ospreys in the Tiran Island (Red Sea) was the blue-
fish (S. cretense) and surgeonfish (Acanthurus spp.) are
spotted cornetfish (Fistularia commersonii; Safriel et
slow-swimming fish that also occur in shallow water
al. 1985), which is a Syngnathiforme broadly similar
along rocky shores, which may make them highly vul-
to A. strigosus which was an important prey on San-
nerable to Ospreys. Among the pelagic species, the
tiago and Sa˜o Vicente (Cape Verde). Likewise, nee-
high consumption of needlefish is likely explained by
dlefish such as Tylosurus spp. were consumed fre-
the habit of this predator of remaining close to the
quently in Cape Verde (Santa Luzia), the Red Sea
water surface, nearly motionless, with the body point-
(Safriel et al. 1985), the Gulf of California (Cartron
ing slightly downwards (Safriel et al. 1985).
and Molles 2002), and the Arabian Gulf (Beech
The estimated mass of fish taken most frequently
2003). In contrast to these species, it is noteworthy
by Ospreys at the Cape Verdes Islands was generally
that mullets (Mugilidade) were very rarely con-
large compared to that in other inland and marine
sumed at Cape Verde, though they are an important
areas (Poole 1989). For example, in Corsica, 94% of
part of Osprey diet along the nearby coast of Sene-
mullet weighed 180–310 g (mean 5 235 g; Francour
gal (Prevost 1982), as they are for many Ospreys
and Thibault 1996), whereas 45.1% of fish prey
feeding along tropical and subtropical coasts (Car-
weighed .300 g in the present study. The greater
tron and Molles 2002, Clancy 2005), and in the
proportion of large fish observed in this study is clos-
Mediterranean (Francour and Thibault 1996). This
er to that recorded in the Red Sea (Safriel et al. 1985)
was probably due to the scarcity of mullets in the
and, to a lesser extent, in New South Wales, Australia
(Clancy 2005). This result suggests that feeding con-
Factors influencing prey selection by Ospreys in
ditions may be particularly favorable in the Cape
Cape Verde are unknown at present, due to the
Verde archipelago, as large prey is generally believed
limited information about coastal fish abundance
to be the most profitable for breeding Ospreys (Glass
and distribution (but see Medina et al. 2007). How-
and Watts 2009). Despite this general pattern, diet
ever, a combination of abundance and susceptibility
on Santiago was largely dominated by A. strigosus,
to capture probably influenced diet composition, as
which is a fish with a long body but with low mass.
found elsewhere (Poole 1989). For instance, cornet-
This, together with the low dietary diversity recorded,
fish may be easily captured because they occur in
may indicate that food resources may be less favor-
shallow reefs and rocky habitats, where they rest,
able on Santiago than elsewhere, which was in agree-
immobile, near the bottom to ambush their prey (Sa-
ment with the small size of the Osprey population
friel et al. 1985). These fish may be particularly abun-
Biplot of a correspondence analysis (CA) of Osprey diet composition in the Cape Verde archipelago (Jan-
uary–June 2006). Species matrix includes species with .50 identified individuals, and combines the remaining prey in an‘‘other fish’’ category. Data from the nearby islets of Raso and Branco were combined due to small sample sizes. Islandsare represented by filled circles; fish species by open squares.
Conservation Implications. A sustainable Osprey
1985, Clancy 2005). We compared the primary spe-
population requires the maintenance of an abun-
cies of demersal fish caught by Cape Verde commer-
dant food supply, as this strongly influences popu-
cial fisheries (Medina et al. 2007) to Osprey diet
lation density and breeding success (Harmata et al.
documented in our study, and found the only over-
2007, Bai et al. 2009, Glass and Watts 2009). In Cape
lap to be A. monroviae, a fish species that contributed
Verde, where large reef and littoral pelagic fish
only 2.1% to the overall Osprey diet. Thus, overfish-
make up the bulk of Osprey diet, maintenance of
ing is unlikely to threaten the population at present,
large stocks of these species will help preserve the
although concentration of fishing effort at the local
Osprey population. One of the potential threats to
scale could still influence prey availability for partic-
Ospreys could thus be overfishing (e.g., Safriel et al.
Estimated mean (6SD) and range of lengths and masses of fish prey species recorded in the diet of Ospreys in
the Cape Verde archipelago (January–June 2006).
Climate change may represent a potentially more
BIRDLIFE INTERNATIONAL. 2010. Species factsheet: Pandion
serious threat for the Osprey in Cape Verde, due to
haliaetus. http://www.birdlife.org (last accessed 20 Sep-
potential reductions in marine productivity and fish
biomass associated with the ongoing moderate
CARTRON, J.-L.E. AND M.C. MOLLES, JR. 2002. Osprey diet
along the eastern side of the Gulf of California, Mexi-
warming of the Canary Current System (Behrenfeld
co. Western North American Naturalist 62:249–252.
et al. 2006, Sherman et al. 2009). Negative effects on
CLANCY, G.P. 2005. The diet of the Osprey (Pandion haliae-
marine productivity may be greater in the Barlavento
tus) on the north coast of New South Wales. Emu
group of islands (Roy and Cury 2003), thereby af-
fecting the Osprey stronghold in the Cape Verde
DUARTE, M.C. AND M.M. ROMEIRAS. 2009. Cape Verde Is-
archipelago (Palma et al. 2004). Thus, there is a
lands. Pages 143–150 in R.G. Gillespie and D.A. Clague
need for long-term monitoring of Osprey popula-
[EDS.], Encyclopedia of islands. University of California
tion densities, breeding success, and diet, to detect
Osprey responses to any changes in food resources.
EINODER, L.D. 2009. A review of the use of seabirds as
Monitoring of Osprey and other generalist piscivo-
indicators in fisheries and ecosystem management.
rous birds could be a relatively simple and inexpen-
sive approach to track temporal and spatial varia-
FLOETER, S.R., L.A. ROCHA, D.R. ROBERTSON, J.C. JOYEUX,
W.F. SMITH-VANIZ, P. WIRTZ, A.J. EDWARDS, J.P. BAR-
tions in littoral fish assemblages across the Cape
REIROS, C.E.L. FERREIRA, J.L. GASPARINI, A. BRITO, J.M.
Verde archipelago. This might provide early warn-
FALCO´N, B.W. BOWEN, AND G. BERNARDI. 2008. Atlantic
ings for changes associated with environmental in-
reef fish biogeography and evolution. Journal of Biogeog-
sults such as overfishing and global warming (e.g.,
FRANCOUR, P. AND J.-C. THIBAULT. 1996. The diet of
breeding Osprey Pandion haliaetus on Corsica: exploita-
tion of a coastal marine environment. Bird Study 43:
This study was partly funded by The International Os-
prey Foundation. We thank Pedro Lo´pez Sua´rez for pro-
FROESE, R. AND D. PAULY. 2010. FishBase. http://www.
viding samples from Boavista, and Augusto Rosa for pro-
fishbase.org (last accessed 8 June 2010).
viding transport to Santa Luzia, Branco and Raso, and
GLASS, K.W. AND B.W. WATTS. 2009. Osprey diet composi-
Guilherme Mascarenhas for spear fishing of cornetfish
tion and quality in high- and low-salinity areas of lower
specimens. We also thank the University of Cape Verde
Chesapeake Bay. Journal of Raptor Research 43:27–36.
for providing the opportunity to undertake this study,and especially the Cape Verdean fishermen from Mindelo,
HARMATA, P.J., M. RESTANI, AND A.R. HARMATA. 2007. Set-
Sa˜o Vicente Island, for helping to collect the material to
tlement patterns, foraging behavior, and reproductive
create the reference collection of fish bones. Useful sug-
success of Ospreys along a heterogeneous riverine cor-
gestions to improve the manuscript were provided by Ka-
ridor. Canadian Journal of Zoology 85:56–62.
ren Steenhof, Chuck Henny, and Rob Bierregaard.
LEGENDRE, P. AND L. LEGENDRE. 1998. Numerical ecology,
Second Ed. Elsevier, Amsterdam, The Netherlands.
LESSICA, P. AND F.W. ALLENDORF. 1995. When are periph-
BAI, M.-L., D. SCHMIDT, E. GOTTSCHALK, AND M. MU¨HLEN-
eral populations valuable for conservation? Conservation
BERG. 2009. Distribution pattern of an expanding Os-
prey (Pandion haliaetus) population in a changing envi-
MEDINA, A., J.-C. BREˆTHES, J.-M. SE´VIGNY, AND B. ZAKARDJIAN.
ronment. Journal of Ornithology 150:255–263.
2007. How geographic distance and depth drive eco-
BANNERMAN, D.A. AND W.N. BANNERMAN. 1968. History of
logical variability and isolation of demersal fish com-
the birds of the Cape Verde Islands. Pages 287–291 in
munities in an archipelago system (Cape Verde, east-
R. Oliver and D. Boyd [EDS.], Birds of the Atlantic
ern Atlantic Ocean). Marine Ecology 28:404–417.
islands, Vol. IV. Oliver and Boyd Ltd., Edinburgh, U.K.
NAUROIS, R. DE. 1987. Le balbuzard (Pandion haliaetus) aux
BEECH, M. 2003. The diet of Osprey Pandion haliaetus on
ıˆles du Cap Vert. Annali del Museo Civico di Storia Natur-
Marawah island, Abu Dhabi Emirate, UAE. Tribulus
ONTIVEROS, D. 2003. Nesting distribution, food habits, and
BEJA, P. 1997. Predation by marine-feeding otters (Lutra
conservation of Osprey on Boavista Island (Archipela-
lutra) in south-west Portugal in relation to fluctuating
go of Cape Verde). Journal of Raptor Research 37:67–70.
food resources. Journal of Zoology 242:503–518.
PALACIOS, C.J. 2004. Current status and distribution of
BEHRENFELD, M.J., R.T. O’MALLEY, D.A. SIEGEL, C.R. MCCLAIN,
birds of prey in the Canary Islands. Bird Conservation
J.L. SARMIENTO, G.C. FELDMAN, A.J. MILLIGAN, P.G. FALK-
OWSKI, R.M. LETELIER, AND E.S. BOSS. 2006. Climate-driven
PALMA, L., J. FERREIRA, R. CANGARATO, AND P.V. PINTO. 2004.
trends in contemporary ocean productivity. Nature
Current status of the Osprey in the Cape Verde Islands.
Journal of Raptor Research 38:141–147.
POOLE, A.F. 1989. Osprey: a natural and unnatural history.
SHERMAN, K., I.M. BELKIN, K.D. FRIEDLAND, J. O’REILLY, AND
Cambridge Univ. Press, New York, NY U.S.A.
K. HYDE. 2009. Accelerated warming and emergent
PREVOST, Y.A. 1982. The wintering ecology of Ospreys in
trends in fisheries biomass yields of the world’s large
Senegambia. Ph.D. dissertation, University of Edin-
marine ecosystems. Ambio 38:215–224.
THIBAULT, J.-C., R. TRIAY, P. BEAUBRUN, D. BOUKHALFA,
REINER, F. 1996. Cata´logo dos peixes do arquipe´lago de
J.M. DOMINICI, AND A. TORRE. 1996. Osprey (Pandion
haliaetus) in the Mediterranean: characteristics of a res-
ident population with a patchy distribution. Pages
ROY, C. AND P. CURY. 2003. Decadal environmental and
135–144 in J. Muntaner and J. Mayol [EDS.], Biologia
ecological changes in the Canary Current Large Marine
y conservacio´n de las rapaces Mediterra´neas. SEO, Ma-
Ecosystem and adjacent waters: patterns of connections
and teleconnection. Pages 255–278 in G. Hempel and
WATTS, B.D. AND B.J. PAXTON. 2007. Ospreys of the Chesa-
K. Sherman [EDS.], Large marine ecosystems of the
peake Bay: population recovery, ecological require-
world: trends in exploitation, protection, and research.
ments, and current threats. Waterbirds 30(Special Pub-
Elsevier, Amsterdam, The Netherlands.
SAFRIEL, U.N., Y. BEN-HUR, AND A. BEN-TUVIA. 1985. The diet
of the Osprey on Tiran Island: management implicationsfor populations on the northern Red Sea islands. Pages181–193 in P.J. Moors [ED.], Conservation of island birds.
Received 8 November 2010; accepted 26 March 2011
ICBP Technical Publication No. 3, Cambridge, U.K.
NeuroImage 12, 707–712 (2000) doi:10.1006/nimg.2000.0660, available online at http://www.idealibrary.com on Abnormal Reactivity of the ϳ20-Hz Motor Cortex Rhythm in UnverrichtLundborg Type Progressive Myoclonus EpilepsyTeija Sile´n,* Nina Forss,* Ole Jensen,* and Riitta Hari*,†* Brain Research Unit, Low Temperature Laboratory, Helsinki University of Technology, P.O. Box 2200, FIN-0
Information for Medicaid Pharmacy Providers and Prescribers Pharmacy Program Changes Effective October 1, 2011 Changes to the Ohio Medicaid Pharmacy program that will be effective on October 1, 2011: 1. Change in pharmacy billing and coverage for members of Medicaid-contracting managed 2. Change to fee-for-service Medicaid Preferred Drug List (PDL) Policy Guidance 1. Change in pha
 Diet of Breeding Ospreys in the Cape Verde Archipelago,
Diet of Breeding Ospreys in the Cape Verde Archipelago,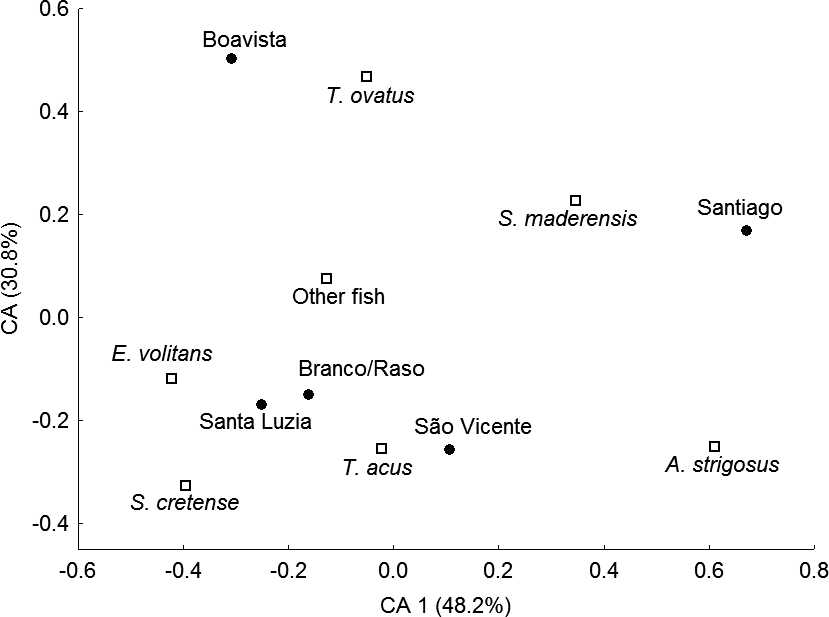 Biplot of a correspondence analysis (CA) of Osprey diet composition in the Cape Verde archipelago (Jan-
uary–June 2006). Species matrix includes species with .50 identified individuals, and combines the remaining prey in an‘‘other fish’’ category. Data from the nearby islets of Raso and Branco were combined due to small sample sizes. Islandsare represented by filled circles; fish species by open squares.
Biplot of a correspondence analysis (CA) of Osprey diet composition in the Cape Verde archipelago (Jan-
uary–June 2006). Species matrix includes species with .50 identified individuals, and combines the remaining prey in an‘‘other fish’’ category. Data from the nearby islets of Raso and Branco were combined due to small sample sizes. Islandsare represented by filled circles; fish species by open squares.