He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Gel.berkeley.edu
Nutrition and Cancer Brown Kelp Modulates Endocrine Hormones in Female Sprague-Dawley Rats and in Human Luteinized Granulosa Cells1
Christine F. Skibola,*2 John D. Curry,*3 Catherine VandeVoort,† Alan Conley,** andMartyn T. Smith*
*School of Public Health, University of California, Berkeley, California; and †California National PrimateResearch Center and **Department of Population Health and Reproduction, School of Veterinary Medicine,University of California, Davis, California
Epidemiological studies suggest that populations consuming typical Asian diets have a lower
incidence of hormone-dependent cancers than populations consuming Western diets. These dietary differenceshave been mainly attributed to higher soy intakes among Asians. However, studies from our laboratory suggest thatthe anti-estrogenic effects of dietary kelp also may contribute to these reduced cancer rates. As a follow-up toprevious findings of endocrine modulation related to kelp ingestion in a pilot study of premenopausal women, we
investigated the endocrine modulating effects of kelp (Fucus vesiculosus) in female rats and human luteinizedgranulosa cells (hLGC). Kelp administration lengthened the rat estrous cycle from 4.3 Ϯ 0.96 to 5.4 Ϯ 1.7 d at 175mg ⅐ kgϪ1 body wt ⅐ dϪ1 (P ϭ 0.05) and to 5.9 Ϯ 1.9 d at 350 mg ⅐ kgϪ1 ⅐ dϪ1 (P ϭ 0.002) and also led to a 100%increase in the length of diestrus (P ϭ 0.02). Following 175 mg ⅐ kgϪ1 ⅐ dϪ1 treatment for 2 wk, serum 17-estradiol
levels were reduced from 48.9 Ϯ 4.5 to 40.2 Ϯ 3.2 ng/L (P ϭ 0.13). After 4 wk, 17-estradiol levels were reducedto 36.7 Ϯ 2.2 ng/L (P ϭ 0.02). In hLGC, 25, 50, and 75 mol/L treatment reduced 17-estradiol levels from 4732
Ϯ 591 to 3632 Ϯ 758, 3313 Ϯ 373, and 3060 Ϯ 538 ng/L, respectively. Kelp treatment also led to modest elevationsin hLGC culture progesterone levels. Kelp extract inhibited the binding of estradiol to estrogen receptor ␣ and  andthat of progesterone to the progesterone receptor, with IC
values of 42.4, 31.8, and 40.7 mol/L, respectively.
These data show endocrine modulating effects of kelp at relevant doses and suggest that dietary kelp maycontribute to the lower incidence of hormone-dependent cancers among the Japanese.
KEY WORDS: ● Fucus vesiculosus ● rat ● seaweed ● breast cancer ● estrogen
The rise in estrogen-dependent cancers in the United
with abnormal menstrual cycling histories (10). In the present
States and our limited success with their prevention and
study, we have further investigated the endocrine modulating
treatment have spurred growing interest in the dietary habits
effects of F. vesiculosus on sex hormone levels and cycling
of the Japanese, who have one of the lowest rates of breast,
patterns in rats and in a human model.
endometrial, and ovarian cancers in the world (1,2). Studies
Previous studies show an inverse relation between men-
show that Japanese women have longer menstrual cycles and
strual cycle length and risk of breast (11), ovarian (12), and
lower serum estradiol levels than their Western counterparts
endometrial (13) cancers. Menstrual cycle length and age of
(3–5), factors that may contribute to their low risk of estrogen-
onset of menarche and menopause may serve as surrogate
dependent cancers. To date, these low rates have been partly
measures of endogenous estradiol and progesterone exposure
attributed to the soy-rich diets inherent among Asian popu-
[reviewed in (14)]. Women with shorter cycles experience a
lations (6 – 8). However, another contributory factor may be
greater total number of menstrual cycles during the course of
their high intake of seaweed, as previously hypothesized by
their reproductive lifetimes than those with longer cycling
Teas et al. (9). In a human pilot study, we demonstrated that
patterns. Hence, these women will spend more time overall in
intake of the brown kelp seaweed, Fucus vesiculosus (bladder-
the follicular and luteal phases of the cycle where estrogen and
wrack), significantly increased the total number of days of the
progesterone levels and endometrial and breast cell prolifera-
menstrual cycle, reduced circulating 17-estradiol levels, and
tion rates are at their highest. Positive associations also have
elevated serum progesterone levels in premenopausal women
been reported among breast, endometria, and ovarian cancersand obesity and alcohol intake, factors that promote persistent
estrogenic stimulation and hormone imbalances (15–17).
Supported by the National Foundation for Cancer Research and NIH Grant
These studies suggest that exposure to estrogens and an im-
2 To whom correspondence should be addressed.
balance in the estrogen/progesterone ratio may be the most
critical determinants in risk of estrogen-dependent diseases.
Present Address: Division of Molecular and Cellular Biology, University of
Inhibition of estrogen via the estrogen receptor or aromatase
0022-3166/05 $8.00 2005 American Society for Nutritional Sciences. Manuscript received 14 September 2004. Initial review completed 27 October 2004. Revision accepted 22 November 2004.
ENDOCRINE MODULATION BY Fucus vesiculosus
blockade is a current strategy for prevention in high-risk
Kelp dose finding experiment (Expt. 1). Normally cycling rats
individuals and in the treatment of some estrogen-dependent
were randomly divided into 3 groups of 8: a vehicle control, a low
diseases. Furthermore, the identification of dietary compo-
dose (175 mg ⅐ kgϪ1 ⅐ dϪ1), and a high dose (350 mg ⅐ kgϪ1 ⅐ dϪ1)
nents that exert chemoprotective effects by suppressing en-
group. Powdered F. vesiculosus was measured and applied in the
dogenous estrogen production may provide another means to
morning daily to a 2-g fresh apple wedge used as a vehicle. There wasa dual advantage to using an apple vehicle in this study: (1) it
reduce the incidence of breast, endometrial, and ovarian can-
eliminated stress associated with gavage; (2) rats eagerly ate the apple
and the kelp in this manner, making it easy to monitor and ensure
Rat and primary human luteinized granulosa cell (hLGC)4
complete deliverance of the kelp. Vaginal smears were obtained and
models have been utilized to examine chemical endocrine
daily logs were maintained to monitor estrous cycling. After ϳ4 wk,
disruptor effects on disease risk. The Sprague-Dawley female
all rats were removed from treatment. No adverse effects were ob-
rat has been used as a model to investigate the effects of
served during the course of the experiment.
endocrine modulation on mammary, ovarian, and endometrial
Time course experiment (Expt. 2). For serum hormone studies
carcinogenesis (18 –20). Chemical endocrine disruptors are
and to determine whether dosing over time altered hormone levels,
commonly tested using the rat model due to similarities in
1-mL blood draws were taken from the tail veins of 19 normally
ovarian hormone responsiveness between humans and rats
cycling rats during the morning of proestrous (determined by vaginal Downloaded from
(21,22). In both the rat estrous cycle and the human menstrual
cytology). Immediately following the blood draw, rats were given 175mg ⅐ kgϪ1 ⅐ dϪ1 kelp. At 2- and 4-wk intervals, blood was redrawn
cycle, estradiol levels peak during proestrus and the follicular
during the morning of proestrus. Blood samples were allowed to clot
phase and progesterone levels peak during diestrus and the
at room temperature and were centrifuged for 10 min at 2000 ϫ g.
luteal phase, respectively. Primary hLGC cultures simulate
Serums were aspirated and frozen at Ϫ20°C until further analysis.
granulosa lutein cells of the corpus luteum in vivo because
High dose experiment (Expt. 3). To determine whether high
they support the timely and dynamic secretion of estradiol and
dose kelp treatments would exert anti-estrogenic and/or progesta- jn.nutrition.org
progesterone in patterns that mimic serum hormone levels
genic effects in rats with high circulating estradiol levels, 8 rats were
during the luteal phase of the menstrual cycle. Thus hLGC
chosen whose estradiol levels were approximately Ն50 g/L. Rats
cultures have been utilized successfully in the study of chem-
were dosed 350 mg ⅐ kgϪ1 ⅐ dϪ1after a baseline blood draw was taken
ical endocrine disruption, such as the effects of dioxin on
during the morning of proestrus. Following 1 wk of treatment, an
ovarian estrogen and progesterone synthesis (22).
additional blood draw was taken during the morning of proestrus.
Our objectives in the present study were to build upon our
Animal hormone assays. Blood serum progesterone was assayed
in triplicate using an ELISA kit (Product No. DSL-10 – 6800) from
previous findings using these models by 1) examining whether
Diagnostic Systems Laboratories according to the manufacturer’s di-
dietary administration of F. vesiculosus disrupts normal estrous
rections. 17-Estradiol was assayed in duplicate by a radioimmune
cycling and sex steroid secretion in Sprague-Dawley rats and
antibody assay according to the method described previously (25). 2) treating hLGC cultures with a F. vesiculosus extract to study
Crude seaweed extractions. Dried, powdered F. vesiculosus (50
differences in endocrine responses in granulosa cells. To fur-
g) was placed in a 1:1 solution of distilled water and 100% ethanol,
ther investigate possible mechanisms of action of the F. ve-
covered, and stirred for 24 h at room temperature. The extract was
siculosus extract in estrogen and progesterone responses, we
centrifuged for 10 min at 4500 ϫ g, sterile filtered, evaporated to
evaluated its binding affinity to estrogen receptor (ER)␣, ER,
dryness using a rotary evaporator, and resolubilized to the desired
and progesterone receptor (PR)-B and its potential to inhibit
concentrations in 50% ethanol. The estimated molecular weight of
aromatase activity in hLGC cultures.
the crude seaweed extract was 300 g/mol, a value commonly used inthe pharmaceutical industry for testing bioactivity of unknown plantcompounds. MATERIALS AND METHODS hLGC culture and treatment. Rats and estrous cycle monitoring.
from 8 women undergoing assisted reproduction treatment at a fer-
Sprague-Dawley rats (Charles River Canada) weighing 200 –250 g
tility clinic. Cells were prepared, plated, and cultured as previously
were individually housed in wire cages. They were allowed ad libitum
described for each patient (26). Briefly, cells were plated in minimum
access to a standard laboratory diet (AIN-76) (23) and water. Fol-
essential medium (MEM) supplemented with 0.1 IU/mL human
lowing a brief 2-wk adjustment period, rats underwent daily vaginal
chorionic gonadotropin, antibiotics, and 5% fetal calf serum at 37°C
cytology monitoring to determine normal estrous cycling using stag-
in an atmosphere of 5% CO in air. After 48 h, cells were treated on
ing criteria described by Everett (24). Rats were required to have at
consecutive days for 9 d with ethanol (vehicle control) or 25, 50, or
least 2 normal, consecutive estrous cycles prior to experimentation. A
75 mmol/L kelp extract. Samples of medium from final day of treat-
normal estrous cycle was defined as a 3- to 5-d cycle. A complete
ment were assayed for 17-estradiol and progesterone.
estrous cycle was defined as the day of estrus to the day before the
Granulosa cell hormone assays. Estradiol and progesterone mea-
subsequent estrus. Weights of rats were monitored weekly throughout
surements were performed using commercially available RIA kits
(Diagnostic Products) as previously reported (27). Source and dosing of F. vesiculosus. Dried, powdered F. vesicu- Estrogen and progesterone receptor binding assays. Affinity of losus was obtained from Maine Coast Sea Vegetables. The kelp was
the kelp extract to ER␣, ER, and PR-B was determined by radio-
harvested from the Gulf of Maine and from the coastal waters of New
metric competitive binding assays as previously described (28,29) by
Brunswick and Nova Scotia during the late summer months. Process-
an outside laboratory (MDS Panlabs). Briefly, dried kelp extract in 3
ing entailed sun drying the entire plant less the holdfast (root sys-
dilutions (0.5, 5, and 50 mol/L final concentration) were resolubi-
tem). The dried seaweed was then milled into a fine powder.
lized in dimethyl sulfoxide, combined with ER␣ or ER and 0.5
Dosage levels were chosen to fall within the range of effective
nmol/L estradiol, and mixed for 2 h at 25°C. Nonspecific binding was
doses in our previous human studies and in a traditional Asian diet,
estimated in the presence of 1 mol/L diethylstilbesterol. To test
taking into account that higher doses are often needed in rats to
PR-B binding, kelp extracts were incubated for 2 h with PR-B and 1.4
produce effects comparable to those seen in humans due to the
nmol/L radiolabeled progesterone at 4°C. Nonspecific binding was
increased rate of metabolism of the rat. Ethical approval was obtained
estimated in the presence of 1 mol/L progesterone. All determina-
for the study and all of the studies were conducted in full compliance
tions were carried out in triplicate and data are means Ϯ SEM.
with the U.C. Berkeley Animal Care and Use Committee guidelines. Aromatase activity measured using a tritiated water assay.
Aromatase activity was estimated by measuring the incorporation oftritium from androstenedione into 3H O as previously described
4 Abbreviations used: ER, estrogen receptor; hLGC, human luteinized gran-
(30,31). Incubations of hLGCs in 500 L MEM with 300 nmol/L
ulosa cell; MEM, minimum essential medium; PR, progesterone receptor.
androstenedione (10% labeled, 90% radio inert, Steraloids) were
carried out at 37°C for 2 h in the presence or absence of the kelpextract (10, 50, and 100 mol/L). Statistical analyses. In Animal Expt. 1, differences between the
means of the 3 groups were evaluated by two-way ANOVA usingProc Mixed in SAS and values are means Ϯ SD. Dunnett’s pairwise
comparison procedure was used to evaluate the pairwise differencesbetween treatments and the control group. For all other experiments,statistical analyses were performed by paired t tests (2-sided) with acommercially available statistical software package (Statsoft) andresults were considered significant for P Ͻ 0.05. Values are means
a nonlinear, least squares regression analysis using Data AnalysisToolbox (MDL Information Systems). Animal studies Kelp dose finding experiment (Animal Expt. 1).
estrous cycle was evaluated daily for ϳ30 d in 24 female
Sprague-Dawley rats. Kelp administration led to a profound,dose-dependent increase in the length of the estrous cycle in
rats fed 175 and 350 mg ⅐ kgϪ1 ⅐ dϪ1 kelp (P ϭ 0.004). In the
17-Estradiol levels in rats following 1 wk of 350
controls, the mean number of days of the estrous cycle was 4.3
mg ⅐ kgϪ1 ⅐ dϪ1 kelp administration. The box plots indicate that in rats
Ϯ 0.96 compared to 5.4 Ϯ 1.7 in the 175 mg ⅐ kgϪ1 ⅐ dϪ1 dose
with high circulating serum 17-estradiol levels median serum 17-
group (P ϭ 0.05) and 5.9 Ϯ 1.9 d in the 350 mg ⅐ kgϪ1 ⅐ dϪ1
estradiol levels significantly decreased from 68.6 to 42.8 ng/L (P
dose group (P ϭ 0.002). Furthermore, kelp treatment led to an
overall 100% increase in the mean length of the diestrus phaseof the estrous cycle (P ϭ 0.02). Specifically, the mean number
of days in diestrus was 0.97 Ϯ 0.22 among the controls com-
extracts compete for and bind to ER␣, ER, and PR-B, with a
pared to 1.4 Ϯ 0.54 for the 175 mg ⅐ kgϪ1 ⅐ dϪ1 dose group and
2.1 Ϯ 0.88 for the 350 mg ⅐ kgϪ1 ⅐ dϪ1 dose group (P ϭ 0.02).
Aromatase activity following treatment of hLGCs with the
Treatment had no significant effect on the number of days in
kelp extract did not differ (data not shown).
estrus, proestrus, or metestrus during the mean estrous cycle. Total number of days monitored was 28.6 Ϯ 3.1, 30.5 Ϯ 3.6,
DISCUSSION
and 31.9 Ϯ 3.6 for the 0, 175, and 350 mg ⅐ kgϪ1 ⅐ dϪ1 groups,
Here we report additional evidence of the anti-estrogenic
Following Expt. 1, 5 rats stopped normal estrous cycling and
bioactivity of dietary F. vesiculosus by demonstrating its effects
were excluded from the remainder of experiments. One re-
on rat estrous cycling patterns and serum hormone levels and
on estradiol production in treated hLGC cultures. Specifically,
Effects on serum estradiol and progesterone levels (Ani-
dietary kelp resulted in an overall 37% increase in the length
mal Expt. 2). Following the 175 mg ⅐ kgϪ1 ⅐ dϪ1 treatment
of the rat estrous cycle in a dose-dependent manner and led to
for 2 wk, mean serum 17-estradiol levels were reduced from
a prolonged diestrus phase of the cycle in the 350
48.9 Ϯ 4.5 to 40.2 Ϯ 3.2 ng/L (P ϭ 0.13) and after 4 wk at the
mg ⅐ kgϪ1 ⅐ dϪ1 group. Kelp administration also exerted a tem-
same dose levels were significantly reduced 25% from baseline
pering effect on estrogen production in rats, which led to
to 36.7 Ϯ 2.2 ng/L (P ϭ 0.02), suggesting an effect of dosing
18 –33% reductions in circulating 17-estradiol levels. These
over time. Serum progesterone levels between controls and the
findings are consistent with the observed increase in menstrual
cycle length and decrease in serum estradiol levels in women
High dose experiment (Animal Expt. 3).
following kelp administration (10). Moreover, the doses used
with high circulating serum 17-estradiol levels following 1
in this study are physiologically relevant doses and are consis-
wk kelp administration (350 mg ⅐ kgϪ1 ⅐ dϪ1), median serum
tent with the range of intakes of 3–13 g/d estimated in Japa-
17-estradiol levels decreased by 38% (P ϭ 0.02) (Fig. 1).
nese populations (32). Previous studies investigating the role
The range in reduction of serum 17-estradiol levels in 6 rats
of dietary soy or genistein on the rat estrous cycle showed
was 25–58%, whereas 2 rats did not respond to kelp at all.
either no effects (33) or only a modest 10% increase in cycle
Progesterone levels were not significantly affected following
length (34), suggesting that kelp may exert a greater effect in
increasing cycle length than soy intake. Effects of kelp treatment on17-estradiol and progester-
The anti-estrogenic bioactivity of F. vesiculosus was further
one levels in human granulosa cells. In hLGC cultures, the
demonstrated in an hLGC bioassay where dosing with kelp
50 and 75 mol/L doses significantly reduced 17-estradiol
extract led to 23–35% reductions in 17-estradiol levels in cell
levels by 30 and 35%, respectively (Table 1). Kelp treatment
cultures. This would suggest that the extract might act by
also led to modest elevations in progesterone in hLGC me-
either inhibiting estradiol production or enhancing its meta-
dium; however, only the 50 mol/L dose was increased (P
bolic breakdown. Competitive inhibition, altered expression,
or posttranslational modification of any one of a number of
Radioligand binding assay.
cytochrome P450 enzymes involved in steroidogenesis (includ-
binding assays, the kelp extract exerted inhibitory effects on
ing cholesterol transport) or in 17-estradiol metabolism
the binding of estradiol to ER␣, ER, and progesterone to
could affect estradiol levels (22,35). However, we found no
PR-B (Table 2). These data demonstrate that F. vesiculosus
inhibitory effects of the kelp extract on aromatase activity,
ENDOCRINE MODULATION BY Fucus vesiculosus
polysaccharides, and the fucosterols found in several brownalgae species. Polyphenols constitute ϳ15% of F. vesiculosus,17-Estradiol and progesterone levels in medium of hLGC
with 25% of this fraction consisting of high-molecular-weight
treated with 0, 25, 50, or 75 mol/L kelp extract1
polymers. Phlorotannins, oligomers, and polymers of phloro-glycinol (1,3,5-trihydroxybenzene) are the largest polyphe-
nolic group found in F. vesiculosus and other brown algae. These compounds have been shown to exert bactericidal ac-
tivity (44), reactive oxygen species inhibition (45), and in-
hibitory effects on human immunodeficiency virus type 1
reverse transcriptase and protease (46). The sulfated polysac-
charides, known as fucoidans, are also found in brown sea-
weeds. Anti-viral (47), anti-proliferative (48), anti-angio-genic, and anti-tumor (49) properties of fucoidans have
1 Values are means Ϯ SEM, n ϭ 4.
recently been described. Because of this complex chemical
2 Based on t tests between the control and the intervention dose.
makeup, the precise nature of the compound(s) responsible for Downloaded from the endocrine-modulating effects described in this study will
which was considered a potential molecular target due to the
be difficult to elucidate but efforts are underway to isolate
fact that a number of plant compounds exert highly specific
inhibitory activity against this enzyme (36 –38). Further stud-
Our data suggest that brown seaweed intake may contribute
ies of the mechanism by which kelp extracts inhibit estradiol
to the lower hormone-dependent cancer rates seen in Asian
production in hLGC cultures are underway.
populations. However, to date we have studied only the F. jn.nutrition.org
Another potential mechanism of endocrine modulation is
vesiculosus species that is found in North America. Whereas
the competitive inhibition of hormone receptors (39). Kelp
brown seaweed is a major constituent of the Asian diet, the
extract served as a competitive inhibitor to the binding of
primary brown seaweeds consumed among Japanese popula-
estradiol to both ER␣ and ER, with a slightly greater selec-
tions include wakame (Unaria pinnatifida) and kombu (Lami-
tivity toward ER than ER␣. These findings suggest that
naria japonica) and not F. vesiculosus. Further studies of the
compounds found in F. vesiculosus could act as estradiol an-
potential endocrine modulating effects of these more com-
tagonists by decreasing the affinity of either ER␣ or ER for its
monly consumed seaweeds are needed along with dietary stud-
ligand. Both receptors, which act as ligand-activated transcrip-
ies of seaweed intake and cancer risk using nutritional epide-
tion factors in target genes, are found in a wide variety of
miology before firm conclusions can be made.
tissues. Despite the relatively similar binding affinities of ER␣
In summary, the detection of dietary components that have
and ER for estradiol, differences in binding specificity be-
estrogen-reducing effects holds promise as a simple means of
tween the ␣ and  ERs and other ligands have been observed
dietary modification to reduce risk of estrogen-dependent can-
(40), although their disparate function in target tissues re-
cers in the general population. Furthermore, the identification
of the anti-estrogenic components in F. vesiculosus may lead to
In the present study, there was no evidence of progestagenic
the discovery of novel selective estrogen receptor modulators
effects of dietary kelp administration in Sprague-Dawley rats,
that may be useful in the treatment and/or prevention of
and only small increases in progesterone were detected in
estrogen-dependent cancers. To this end, the isolation and
hLGC cultures following treatment with the kelp extract. Yet,
identification of active components are currently in progress.
it was previously shown that F. vesiculosus administrationcorrected a progesterone deficiency in a woman with a lutealphase defect exhibiting high circulating estradiol levels (10). These discrepancies may be due to the abnormally high estra-
diol-to-progesterone ratio in the subject previously studied. Percentage inhibition, IC50 and Ki values of kelp extracts
There is evidence that estradiol, at relatively high but physi-
(0.5, 5, and 50 mol/L) on the binding of radioligands
ologic concentrations, is a direct inhibitor of 3-hydroxy-
steroid dehydrogenase, resulting in progesterone suppression in
to their respective receptors (ER␣, ER, and PR-B)
human luteal cells (41,42). Thus, in those with high circulat-
in competitive radioligand binding assays
ing estradiol, the anti-estrogenic activity of F. vesiculosus maynot only abrogate estradiol production, but also enhance pro-
gesterone formation by alleviating estradiol’s inhibitory effects
on 3-hydroxysteroid dehydrogenase. However, the potential
impact of dietary kelp on circulating progesterone levels needsto be studied in a larger population.
One limitation of this study is that all rat blood was drawn
during the morning of proestrus when estradiol levels were
highest, but progesterone levels were not at their peak. Due to
multiple blood draws from each rat, we were unable to obtain
blood during other phases of the rat estrous cycle. However, we
were able to determine the relative binding affinity of the kelp
extract for PR-B, suggesting that kelp may act as a PR agonist.
Chemical analysis of F. vesiculosus has revealed that it
contains many potentially bioactive compounds (43). Cur-rently, we are in the process of identifying which agents in F.vesiculosus are responsible for its anti-estrogenic activity. Pos-
sible candidates include the bioactive polyphenols, sulfated
3 Negative values correspond to stimulation of binding. ACKNOWLEDGMENTS
hormone (GnRH) induced luteinizing hormone (LH) secretion from perfusedequine pituitaries. Domest. Anim. Endocrinol. 8: 353–368.
We thank Satyabrata Nandi and Raphael Guzman for providing
Breast cancer and hormone-replacement therapy in
training and technical expertise in estrous cycling cytology determi-
the Million Women Study. Lancet 362: 419 – 427.
nation. We also thank Howard Bern of U.C. Berkeley for his guid-
27. Enan, E., Lasley, B., Stewart, D., Overstreet, J. & Vandevoort, C. A.
2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) modulates function of hu-
ance in the design of our animal studies.
man luteinizing granulosa cells via cAMP signaling and early reduction of glucosetransporting activity. Reprod. Toxicol. 10: 191–198. LITERATURE CITED
28. Carbajo, P., Christensen, K., Edwards, D. P. & Skafar, D. F.
Binding of [3H]progesterone to the human progesterone receptor: differences
1. Parkin, D. M., Pisani, P. & Ferlay, J.
between individual and mixed isoforms. Endocrinology 137: 2339 –2346.
incidence of 25 major cancers in 1990. Int. J. Cancer 80: 827– 841.
29. Obourn, J. D., Koszewski, N. J. & Notides, A. C.
2. Parkin, D. M., Pisani, P. & Ferlay, J.
DNA-binding mechanisms of the recombinant human estrogen receptor. Bio-
3. Olsson, H., Landin-Olsson, M. & Gullberg, B.
30. Corbin, C. J., Trant, J. M., Walters, K. W. & Conley, A. J.
assessment of menstrual cycle length in patients with breast cancer, in patients
Changes in testosterone metabolism associated with the evolution of placental
with benign breast disease, and in women without breast disease. J. Natl. Cancer
and gonadal isozymes of porcine aromatase cytochrome P450. Endocrinology
4. Shimizu, H., Ross, R. K., Bernstein, L., Pike, M. C. & Henderson, B. E.
31. Lephart, E. D. & Simpson, E. R.
Assay of aromatase activity. Downloaded from
Serum oestrogen levels in postmenopausal women: comparison of
American whites and Japanese in Japan. Br. J. Cancer 62: 451– 453.
32. Teas, J., Hebert, J. R., Fitton, J. H. & Zimba, P. V.
5. Key, T. J., Chen, J., Wang, D. Y., Pike, M. C. & Boreham, J.
poor man’s HAART? Med. Hypotheses 62: 507–510.
hormones in women in rural China and in Britain. Br. J. Cancer 62: 631– 636.
33. Masutomi, N., Shibutani, M., Takagi, H., Uneyama, C., Takahashi, N. &
6. Lu, L. J., Anderson, K. E., Grady, J. J., Kohen, F. & Nagamani, M.
Impact of dietary exposure to methoxychlor, genistein, or
Decreased ovarian hormones during a soya diet: implications for breast cancer
diisononyl phthalate during the perinatal period on the development of the rat
prevention. Cancer Res. 60: 4112– 4121.
endocrine/reproductive systems in later life. Toxicology 192: 149 –170.
7. Setchell, K. D., Borriello, S. P., Hulme, P., Kirk, D. N. & Axelson, M.
34. Gallo, D., Cantelmo, F., Distefano, M., Ferlini, C., Zannoni, G. F., Riva, A., jn.nutrition.org
Nonsteroidal estrogens of dietary origin: possible roles in hormone-
Morazzoni, P., Bombardelli, E., Mancuso, S. & Scambia, G.
dependent disease. Am. J. Clin. Nutr. 40: 569 –578.
tive effects of dietary soy in female Wistar rats. Food Chem. Toxicol. 37: 493–502.
8. Cassidy, A., Bingham, S. & Setchell, K. D.
35. Zhu, B. T., Lech, J., Rosen, R. T. & Conney, A. H.
a diet of soy protein rich in isoflavones on the menstrual cycle of premenopausal
dietary 2(3)-tert-butyl-4-hydroxyanisole on the metabolism and action of estradiol
women. Am. J. Clin. Nutr. 60: 333–340.
and estrone in female CD-1 mice. Cancer Res. 57: 2419 –2427.
9. Teas, J., Harbison, M. L. & Gelman, R. S.
36. Le Bail, J. C., Laroche, T., Marre-Fournier, F. & Habrioux, G.
(Laminaria) and mammary carcinogenesis in rats. Cancer Res. 44: 2758 –2761.
Aromatase and 17beta-hydroxysteroid dehydrogenase inhibition by flavonoids.
A pilot study into the effect of intake of Fucus
vesiculosus, an edible brown seaweed, upon menstrual cycle length and hor-
37. Pouget, C., Fagnere, C., Basly, J. P., Besson, A. E., Champavier, Y.,
monal status in pre-menopausal women. Biomed. Central. 4: 10.
Synthesis and aromatase inhibitory activity
11. Beiler, J. S., Zhu, K., Hunter, S., Payne-Wilks, K., Roland, C. L. & Chin-
of flavanones. Pharm. Res. 19: 286 –291.
A case-control study of menstrual factors in relation to
38. Eng, E. T., Ye, J., Williams, D., Phung, S., Moore, R. E., Young, M. K.,
breast cancer risk in African-American women. J. Natl. Med. Assoc. 95: 930 –938.
Gruntmanis, U., Braunstein, G. & Chen, S.
12. Purdie, D. M., Bain, C. J., Siskind, V., Webb, P. M. & Green, A. C.
biosynthesis by procyanidin dimers in red wine and grape seeds. Cancer Res. 63:
Ovulation and risk of epithelial ovarian cancer. Int. J. Cancer 104: 228 –232.
13. Xu, W. H., Xiang, Y. B., Ruan, Z. X., Zheng, W., Cheng, J. R., Dai, Q., Gao,
A review of selective estrogen receptor modulators in
Menstrual and reproductive factors and endometrial
the treatment of breast and endometrial cancer. Semin. Oncol. 29: 129 –133.
cancer risk: Results from a population-based case-control study in urban Shang-
40. Harris, H. A., Bapat, A. R., Gonder, D. S. & Frail, D. E.
hai. Int. J. Cancer 108: 613– 619.
binding profiles of estrogen receptors alpha and beta are species dependent.
14. Kelsey, J. L., Gammon, M. D. & John, E. M.
factors and breast cancer. Epidemiol. Rev. 15: 36 – 47.
41. Vega, M., Devoto, L., Castro, O. & Kohen, P.
15. Madigan, M. P., Troisi, R., Potischman, N., Dorgan, J. F., Brinton, L. A. &
synthesis by human luteal cells: modulation by estradiol. J. Clin. Endocrinol.
Serum hormone levels in relation to reproductive and
lifestyle factors in postmenopausal women (United States). Cancer Causes Con-
42. Fisch, B., Rose, M. P., Elder, M. G., Winston, R. M., Margara, R. A. &
Effects of oestrogen on progesterone synthesis and
Behavioral risk factors in breast cancer: can risk
arachidonic acid metabolism in human luteal cells. Clin. Endocrinol. (Oxf.) 40:
be modified? Oncologist 8: 326 –334.
17. Tavani, A., Bosetti, C., Dal Maso, L., Giordano, L., Franceschi, S. & La
43. Ragan, M. A. & Craigie, J. S.
Influence of selected hormonal and lifestyle factors on
compounds of brown algae. Isolation and characterization of phloroglucinol poly-
familial propensity to ovarian cancer. Gynecol. Oncol. 92: 922–926.
18. Schedin, P., Mitrenga, T. & Kaeck, M.
mers from Fucus vesiculosus (L.). Can. J. Biochem. 54: 66 –73.
of mammary epithelial cell proliferation, differentiation, and death in the Sprague-
44. Nagayama, K., Iwamura, Y., Shibata, T., Hirayama, I. & Nakamura, T.
Dawley rat: a model for investigating the role of estrous cycling in mammary
Bactericidal activity of phlorotannins from the brown alga Ecklonia ku-
carcinogenesis. J. Mammary Gland Biol. Neoplasia 5: 211–225.
rome. J. Antimicrob. Chemother. 50: 889 – 893.
19. Stewart, S. L., Querec, T. D., Gruver, B. N., O’Hare, B., Babb, J. S. &
45. Kang, H. S., Chung, H. Y., Kim, J. Y., Son, B. W., Jung, H. A. & Choi, J. S.
Gonadotropin and steroid hormones stimulate proliferation
Inhibitory phlorotannins from the edible brown alga Ecklonia stolonifera
of the rat ovarian surface epithelium. J. Cell Physiol. 198: 119 –124.
on total reactive oxygen species (ROS) generation. Arch. Pharm. Res. 27: 194 –
20. Poulet, F. M., Roessler, M. L. & Vancutsem, P. M.
alterations caused by developmental exposure to tamoxifen. Reprod. Toxicol. 11:
46. Ahn, M. J., Yoon, K. D., Min, S. Y., Lee, J. S., Kim, J. H., Kim, T. G., Kim,
S. H., Kim, N. G., Huh, H. & Kim, J.
21. Safe, S., Wang, F., Porter, W., Duan, R. & McDougal, A.
transcriptase and protease by phlorotannins from the brown alga Ecklonia cava.
receptor agonists as endocrine disruptors: antiestrogenic activity and mecha-
nisms. Toxicol. Lett. 102–103: 343–347.
47. Ponce, N. M., Pujol, C. A., Damonte, E. B., Flores, M. L. & Stortz, C. A.
22. Moran, F. M., Conley, A. J., Corbin, C. J., Enan, E., VandeVoort, C.,
Fucoidans from the brown seaweed Adenocystis utricularis: extraction
Overstreet, J. W. & Lasley, B. L.
2,3,7,8-Tetrachlorodibenzo-p-dioxin
methods, antiviral activity and structural studies. Carbohydr. Res. 338: 153–165.
decreases estradiol production without altering the enzyme activity of cyto-
48. Funahashi, H., Imai, T., Tanaka, Y., Tsukamura, K., Hayakawa, Y., Kiku-
chrome P450 aromatase of human luteinized granulosa cells in vitro. Biol. Reprod.
mori, T., Mase, T., Itoh, T., Nishikawa, M., Hayashi, H., Shibata, A., Hibi, Y.,
ation of 7,12-dimethylbenz(a)-anthracene-induced mammary tumors in rats. Jpn.
Standards for Nutritional Studies. J. Nutr. 110: 1717–1726.
Neurobiology of reproduction in the female rat. A
49. Koyanagi, S., Tanigawa, N., Nakagawa, H., Soeda, S. & Shimeno, H.
fifty-year perspective. Monogr. Endocrinol. 32: 1–133.
Oversulfation of fucoidan enhances its anti-angiogenic and antitumor
25. Pinaud, M. A., Roser, J. F. & Dybdal, N.
activities. Biochem. Pharmacol. 65: 173–179.
JPP 2005, 57: 1–6 ß 2005 The AuthorsReceived September 23, 2004Accepted February 2, 2005Nelumbinis Semen reverses a decrease in hippocampal5-HT release induced by chronic mild stress in ratsMoonkyu Kang, Kwang-Ho Pyun, Choon-Gon Jang, Hyuntaek Kim,Depression is associated with a dysfunctional serotonin system. Recently, several lines of evidencehave suggested that a very important evo
T e c h n i s c h e s M e r k b l a t t SIGMA Clean SG Beschreibung: SIGMA Clean SG ist eine seidenglänzende, robuste, hoch strapazierfähige und sehr gut reinigungsfähige Beschichtung für beanspruchte Oberflächen im Innenbereich. Verwendungszweck: Besonders geeignet für Beschichtungsarbeiten an Wand- und Deckenflächen mit seidenglänzendem Oberflächen-Finish an die
 Nutrition and Cancer
Nutrition and Cancer ENDOCRINE MODULATION BY Fucus vesiculosus
blockade is a current strategy for prevention in high-risk
Kelp dose finding experiment (Expt. 1). Normally cycling rats
ENDOCRINE MODULATION BY Fucus vesiculosus
blockade is a current strategy for prevention in high-risk
Kelp dose finding experiment (Expt. 1). Normally cycling rats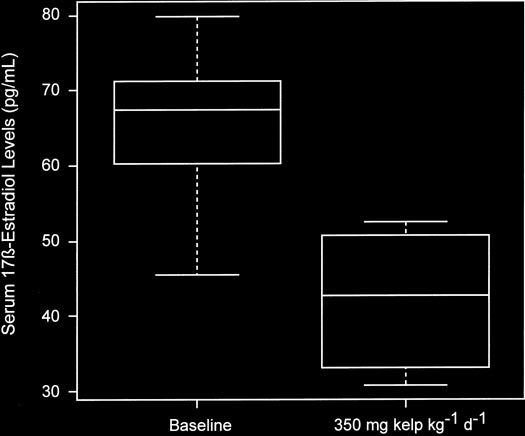
 carried out at 37°C for 2 h in the presence or absence of the kelpextract (10, 50, and 100 mol/L).
carried out at 37°C for 2 h in the presence or absence of the kelpextract (10, 50, and 100 mol/L). ENDOCRINE MODULATION BY Fucus vesiculosus
polysaccharides, and the fucosterols found in several brownalgae species. Polyphenols constitute ϳ15% of F. vesiculosus,
17-Estradiol and progesterone levels in medium of hLGC
with 25% of this fraction consisting of high-molecular-weight
treated with 0, 25, 50, or 75 mol/L kelp extract1
polymers. Phlorotannins, oligomers, and polymers of phloro-glycinol (1,3,5-trihydroxybenzene) are the largest polyphe-
nolic group found in F. vesiculosus and other brown algae.
ENDOCRINE MODULATION BY Fucus vesiculosus
polysaccharides, and the fucosterols found in several brownalgae species. Polyphenols constitute ϳ15% of F. vesiculosus,
17-Estradiol and progesterone levels in medium of hLGC
with 25% of this fraction consisting of high-molecular-weight
treated with 0, 25, 50, or 75 mol/L kelp extract1
polymers. Phlorotannins, oligomers, and polymers of phloro-glycinol (1,3,5-trihydroxybenzene) are the largest polyphe-
nolic group found in F. vesiculosus and other brown algae. ACKNOWLEDGMENTS
ACKNOWLEDGMENTS