He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Bcm_1043.qxd
ISSN 0006-2979, Biochemistry (Moscow), 2013, Vol. 78, No. 9, pp. 1043-1047. Pleiades Publishing, Ltd., 2013. Original Russian Text F. F. Severin, B. A. Feniouk, V. P. Skulachev, 2013, published in Biokhimiya, 2013, Vol. 78, No. 9, pp. 1331-1336.Advanced Glycation of Cellular Proteins as a Possible Basic Component of the “Master Biological Clock” F. F. Severin1,2, B. A. Feniouk2,3, and V. P. Skulachev1,2,3* 1Lomonosov Moscow State University, Belozersky Institute of Physico-Chemical Biology,119991 Moscow, Russia; E-mail: [email protected]2Lomonosov Moscow State University, Institute of Mitoengineering, 119992 Moscow, Russia3Lomonosov Moscow State University, Faculty of Bioengineering and Bioinformatics, 119991 Moscow, RussiaAbstract—During the last decade, evidence has been accumulating supporting the hypothesis that aging is genetically pro- grammed and, therefore, precisely timed. This hypothesis poses a question: what is the mechanism of the biological clock that controls aging? Measuring the level of the advanced glycation end products (AGE) is one of the possible principles underlying the functioning of the biological clock. Protein glycation is an irreversible, non-enzymatic, and relatively slow process. Moreover, many types of cells have receptors that can measure AGE level. We propose the existence of a protein that has a lifespan comparable to that of the whole organism. Interaction of the advanced glycation end product generated from this protein with a specific AGE receptor might initiate apoptosis in a vitally important non-regenerating tissue that produces a primary juvenile hormone. This could result in the age-dependent decrease in the level of this hormone leading to aging of the organism. DOI: 10.1134/S0006297913090101 Key words: biological clock, aging, protein glycation, melatonin, juvenile hormone, AGE, RAGE
Within the phenoptosis concept, aging is considered
organisms in general. However, there are other sponta-
to be the last stage of ontogenesis, the individual develop-
neous chemical processes that might be the basis of the
ment of an organism [1-4]. However, if this is the case,
mechanism measuring many years. L→D isomerization of
then both aging and ontogenesis as a whole should be
amino acids in long-lived proteins is an example of this
managed by a “master biological clock” as postulated by
type of process. In whales, crystallins (lens proteins whose
V. M. Dilman [5, 6] and later by A. Comfort [7]. Our
age is comparable to the maximum age of the animal, i.e.
knowledge of the molecular mechanism of the measure-
about two centuries) have been described [8]. Initially, all
ment of time by living organisms has so far been limited
the amino acids in crystallin are L-isomers. Over many
to circadian rhythm. This rhythm is determined by cyclic
years, the L-amino acids spontaneously isomerize into D-
biochemical reactions that take place in the epiphysis (in
isomers. This process is the fastest for aspartate (about 2%
birds) or in the suprachiasmatic nucleus of the hypothal-
in 10 years). This means that in case of a 200-year-old
amus (in mammals). The signals formed by these organs
whale, there are 40% aspartate residues that are present as
cause fluctuations in concentration of certain hormones
D-isomers in crystallin. A counter of biological age might
in the blood, especially of melatonin. It seems obvious
be formed from a protein similar to crystallin and some
that measurement of time on the scale of hours inherent
device that would measure D-aspartate content in the pro-
to the circadian mechanism is hardly suitable for time
tein. Humans have a number of other proteins (besides
measurement on the scale of decades.
crystallin) that are formed only once in a lifetime. Such
Humans check time by comparing it with a reference
proteins have been found in tooth enamel, white matter of
device based on measuring the rate of radioactive decay. It
the brain, aorta, arteries, skin, cartilage, bones, and ten-
is unlikely that such a mechanism could be used by living
dons. Elastin is also a protein of this type [9].
Asparagine and glutamine deamidation in proteins can
also be spontaneous. The rate of this process has been shown
Abbreviations: AGE, Advanced Glycation End products; GRH,
to depend on the protein conformation. It seems quite inter-
gonadotropin-releasing hormone; mROS, mitochondrial reac-tive oxygen species; RAGE, receptors of AGE.
esting that the rate of spontaneous deamidation varies from
* To whom correspondence should be addressed.
several hours to hundreds of years. N. I. and A. B. Robinson,
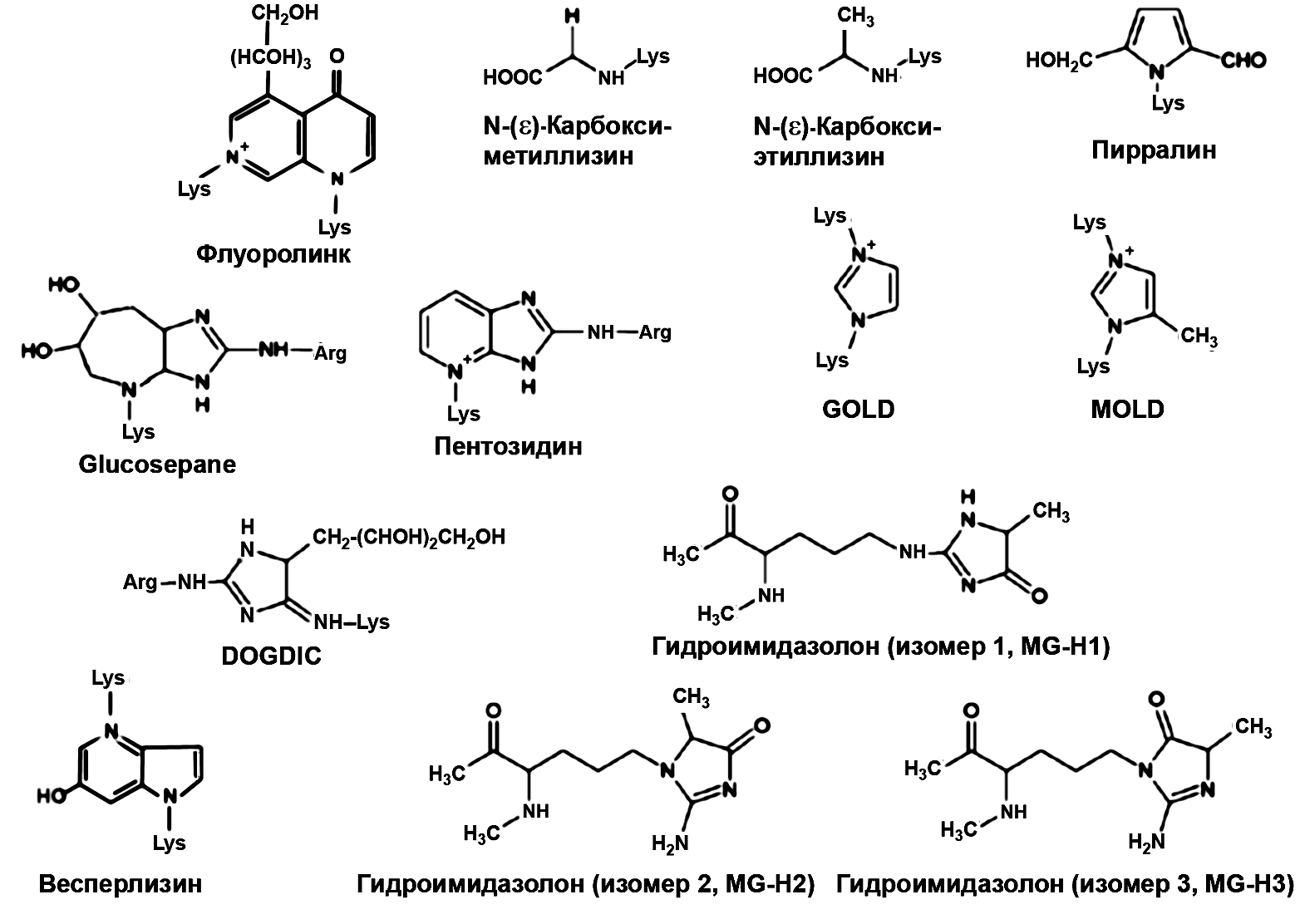
N-e-carboxy- N-e-carboxy- methyl-lysine ethyl-lysine Pyrroline Fluorolink Pentosidine Hydroimidazolone (isomer 1, MG-H1) Vesperlysine Hydroimidazolone (isomer 2, MG-H2) Hydroimidazolone (isomer 3, MG-H3) Fig. 1. Chemical formulas of the most common AGE [13, 14].
who published a number of articles [10-12] and a special
reducing sugars mixed with peptides and proteins. The
book [11a] devoted to the description of this phenomenon
“Maillard reaction”, being a redox process, is a complex
believe some “molecular clock” measuring the time of the
network of successive and parallel reactions.
protein’s life to be hidden in its amino acid sequence.
John E. Hodge [16] contributed greatly to the under-
The process of accumulation of so-called AGE
standing of the chemistry of the Maillard reaction. He
(Advanced Glycation End products) is another candidate
showed that the sugar aldehyde group reacts with amino
for the role of the key component of the “master biological
groups producing N-glycosides at the initial stage of gly-
clock”. AGE embrace a group of diverse compounds that
cation (non-enzymatic glycosylation). Then N-glyco-
are formed in an organism as a result of complex chemical
sides undergo Amadori rearrangement, turning into 1-
reactions, glycation of proteins and peptides being the first
amino-1-deoxy-ketoses (Amadori compounds). These
stage of this process (Fig. 1). These chemical reactions pro-
reactions are followed by further chemical transforma-
ceed slowly but inevitably in any aqueous solution contain-
tions resulting in the formation of AGE [16].
ing proteins and reducing sugars. They result in the forma-
Dicarbonyl products (glyoxal and methylglyoxal),
tion of characteristic colored products as well as covalent,
formed as intermediate products in the course of the
enzymatically uncleavable cross-links in proteins.
Maillard reaction, are of great importance. These highlyactive compounds, which are also formed in the cell asglycolysis byproducts, can react with proteins to form
enzymatically uncleavable cross-links.
Comparison of various amino acid residues in pep-
In 1912, Louis Camille Maillard reported that
tides according to their ability to react with reducing sug-
reducing sugars react with amino acids in solution pro-
ars revealed that side chains of cysteine, lysine, and histi-
ducing dark-colored products (melanoidins) [15]. Similar
dine, as well as the amino group of the N-terminal amino
chemical reactions could be observed also in solutions of
acid, have the highest relative activity. In the case of pep-
AGE AND THE “MASTER BIOLOGICAL CLOCK”
tides reacting with protein AGE-products, the highest
oligosaccharyl transferase-48 (AGE-R1), phosphoprotein
efficiency of cross-link formation was observed for argi-
80K-H (AGE-R2), and galectin-3 (AGE-R3) [29].
AGE receptors were originally assumed to bind and
neutralize AGE. However, further studies showed AGEbinding to RAGE receptor of macrophages and
microglial cells triggers oxidative stress and causes activa-
tion of the p21ras/MAP-kinase signaling cascade, whichin turn activates NF-κB factor [30]. As a result, the inter-
Just as increased concentration of reactive oxygen
action of AGE with their receptors causes inflammatory
species (ROS) causes oxidative stress, increased concen-
response and oxidative stress. In this regard, the role of
tration of sugars (glucose, fructose, deoxyglucose, and
AGE as a biological clock is very likely to be realized via
triose phosphates) and active dicarbonyl compounds (gly-
inflammatory response, ROS generation, and apoptosis
oxal and methylglyoxal) can cause “carbonyl stress”
resulting in the increased rate of formation of AGE prod-
For such a clock to control aging, the following com-
ucts, including cross links in proteins, which violate their
ponents are needed: 1) a protein like crystallin or elastin
whose lifetime is comparable to the human lifespan; 2)
In the early 1980s, after AGE had been found to
RAGE specific to the formation of AGE in this protein;
accumulate with age in certain tissues of living organisms,
3) an apoptosis inducer triggered by the complex of
a theory of “non-enzymatic glycosylation as the cause of
RAGE and an AGE-containing protein. If these process-
es take place in tissue producing some primary juvenile
Harmful effects of glycation were initially assumed to
hormone, and the ability this tissue for restoration is lim-
be associated with damage to long-lived proteins. Indeed,
ited, it should result in the involution of the tissue with
clear age-dependent increase in AGE concentration is
age and subsequent aging due to reduction of the level of
observed in those body parts where proteins are not
renewed or are renewed very slowly (e.g. in skin and car-
This scheme assumes such long time periods as
tilage collagen [19-21], in proteins of the lens [22, 23],
months and years are measured not by a single cell, but by
and in some other tissues [24, 25]). In experiments on
a large group of cells, a special supracellular structure sim-
rats, aminoguanidine, an inhibitor of AGE formation,
ilar to the suprachiasmatic nucleus of the hypothalamus or
was shown to reduce manifestation of such signs of aging
the epiphysis producing melatonin, the hormone of circa-
as decrease in elasticity of blood vessels and increase in
dian rhythm. It seems interesting that the level of mela-
heart size [26]. Furthermore, an important role of AGE
tonin, antioxidant and inducer of the entire group of
was demonstrated in the development of atherosclerosis,
enzymes of the cell antioxidant system, drastically
cataract, diabetic nephropathy, retinopathy, neuropathy,
decreases with age. Furthermore, melatonin has a gero-
and age-dependent vascular complications [13].
protective effect [31-34]. In other words, there is a possi-
Further research showed a significant proportion of
bility that circadian rhythm and general age are measured
the harmful effects of AGE accumulation to be associat-
by the same organ. Reduction of the number of cells pro-
ed with certain signals produced by RAGE (Receptor for
ducing primary juvenile hormone might result in decrease
Advanced Glycation End products) – a specific AGE
in its level in blood and other tissues of the organism. We
receptor, as well as reactive oxygen species inducer. Thus,
believe this could serve as a signal for increasing the con-
although the kinetics of glycation may be similar to kinet-
centration of ROS in mitochondria (mROS). This, in turn,
ics of, for example, isomerization of amino acids in pro-
stimulates apoptosis and causes reduction in the number of
teins (AGE level in elastin of the intervertebral discs
cells in these tissues, which results in deterioration of the
increases with age in approximately the same proportion
physiological functions of the organism [4, 35, 36] (Fig. 2).
as the amount of D-aspartate [9]), the presence of AGE
It should be emphasized that the primary juvenile
receptors makes them a far more likely candidate for the
hormone is likely to trigger a hormonal cascade com-
posed of secondary, tertiary, and other juvenile hormones,which multiply the signal of the primary hormone andtransmit this signal to other organs and tissues. Very
recently, a mechanism controlling aging by one such sec-ondary juvenile hormone, namely, gonadotropin-releas-
RAGE is to be the most thoroughly-studied AGE
ing hormone (GRH), has been discovered. In the group
receptor. It is a multi-ligand membrane receptor of the
of the American cell biologist D. Cai [37], aging was
immunoglobulin superfamily present in many cell types [27,
shown to be accompanied by an increase in the number of
28]. In addition to RAGE, several other receptors binding
microglial cells (playing the role of phagocytes in brain)
AGE products have been discovered, including phagocytic
of one of the sections of hypothalamus and by activation
receptors of macrophages (of the first and second types),
of transcription factor NF-κB in these cells. NF-κB
is maintained mainly at the same level during the day
(increases in its level following the consumption of food
rich in glucose are relatively short-lived). However, amongall the natural monosaccharides, glucose is characterized
by equilibrium of aldehyde and cyclic isoforms, being
maximally shifted in the direction of the cyclic isoform
(only 0.2% of glucose is in the form of aldehyde capable of
being an AGE precursor and, therefore, of participating in
glycation of proteins). Thus, glucose is one of the least
of the “master biological clock” organ
active sugars in relation to glycation. It seems very likely
that glucose became the main “carbohydrate energy carri-er” during evolution exactly due to this property [13]. On
Reduction in the level of juvenile hormone
the other hand, the ability of the above-mentioned
byproduct of glucose metabolism, methylglyoxal, to form
AGE products exceeds the reactive ability of glucose 650-
fold (in the reaction with β-alanine at 80oC) [38].
In the case of galactose, the amount of aldehyde form
many times exceeds that of glucose [39]. Glycation ofcrystallins and serum albumin at the expense of galactose
is much faster than that at the expense of glucose and fruc-
tose [40, 41]. Moreover, addition of galactose to the diet
was shown to cause typical progeria (premature aging)
[42-44], in which the mitochondrial path of apoptosis,
when cytochrome c is released from mitochondria, playsan important role [45]. Development is progeria inhibited
Fig. 2. Hypothetical mechanism of the “master biological clock”
by salidroside, an inhibitor of RAGE-type receptors [46].
controlling ontogenesis in its last stage, the aging of the organism.
Metformin, inhibiting AGE formation from monosaccha-rides [47], is also known to be a geroprotector [48].
RAGE can be found in many cell types, in particular,
causes microglia to produce tumor necrosis factor
in proinflammatory ones [27, 28]. According to in vitro
(TNF), which attacks neighboring neurons responsible
data, not only AGE, but also a number of marker mole-
for GRH synthesis. Neurons possess their own NF-κB
cules and activators of inflammatory processes can be
activated by TNF. This activation in turn stimulates
RAGE ligands: DNA and RNA, calcium-binding pro-
methylation of the GRH gene promotor, which blocks
teins of the S100 family, prions, and the non-histone
GRH synthesis by neurons. Without GRH, the hypo-
chromosomal protein HMGB1, which together with
physis does not produce a tertiary juvenile hormone
RAGE play an important role in inflammatory (including
(gonadotropin) that required for the synthesis of sex hor-
septic and autoimmune) reactions and cancers [28, 49,
mones and a number of systems operating in a young
50]. In addition, some RAGE bind β-amyloid, a protein
organism and weakening with age. By blocking the above-
playing the key role in Alzheimer’s disease [27-29]. Also,
described regulatory chain, the authors managed to pro-
RAGE knockout was shown to promote survival of ani-
long the life of mice and inhibit the development of such
mals with diabetes [51] or bacterial infection [28]. That
signs of aging as sarcopenia, osteoporosis, skin thinning,
would suggest diabetes (violation of sugar metabolism),
appearance of cross links in tendon tissue, and memory
inflammatory diseases, and Alzheimer’s disease to speed
loss. Partial inhibition of the development of these symp-
toms could be achieved by subcutaneous administration
Finally, having accepted the hypothesis of the “mas-
of GRH to old mice. Within the scheme (Fig. 2), the
ter biological clock”, we can put forward the question of
events described by Cai et al. take place between the pri-
the scope of its application. It is natural to assume that
mary juvenile hormone and mROS in the cells of organs
such a clock might be used not only in aging, but also in
and tissues. It is noteworthy that neurons forming the sec-
the earlier stages of ontogenesis. This assumption is con-
ondary juvenile hormone GRH are localized in the hypo-
sistent with the data on the participation of RAGE in dif-
thalamus, i.e. the same place as the suprachiasmatic
nucleus with its “clock” of circadian rhythm.
Thus, results of many experiments in the fields of
Considering the scheme in Fig. 2, we can ask a ques-
gerontology, developmental biology, and medicine can be
tion: which sugar is the source of AGE if these compounds
proposed using the hypothesis of an AGE-based “biolog-
are actually used for measuring age? Glucose seems to be
ical clock”. Therefore, there are good reasons to believe
a good candidate for this role as its concentration in blood
that this hypothesis will be tested in the near future.
AGE AND THE “MASTER BIOLOGICAL CLOCK”
Note added in proof: After this manuscript was accept-
24. Schleicher, E. D., Wagner, E., and Nerlich, A. G. (1997) J.
ed for publication, the paper of C. Menni et al. (C. Menni et
Clin. Invest., 99, 457-468.
al. (2013) Int. J. Epidemiol., July 8, doi: 10.1093/ije/dyt094)
25. Simm, A., Wagner, J., Gursinsky, T., Nass, N., Friedrich,
appeared. The authors showed that blood level of one of the
I., Schinzel, R., Czeslik, E., Silber, R. E., and Scheubel, R. J. (2007) Exp. Gerontol., 42, 668-675.
protein glycation products, i.e. C-glycosyl tryprophan, cor-
26. Corman, B., Duriez, M., Poitevin, P., Heudes, D.,
relates with age and such age-related parameters as lung
Bruneval, P., Tedgui, A., and Levy, B. I. (1998) Proc. Natl.
functional state and bone mineral density. The investigation
Acad. Sci. USA, 95, 1301-1306.
was carried out on humans (6055 persons).
27. Basta, G. (2008) Atherosclerosis, 196, 9-21. 28. Ramsgaard, L., Englert, J. M., Manni, M. L., Milutinovic,
P. S., Gefter, J., Tobolewski, J., Crum, L., Coudriet, G. M.,
Piganelli, J., Zamora, R., Vodovotz, Y., Enghild, J. J., and Oury, T. D. (2011) PLoS One, 6, e20132.
1. Skulachev, V. P. (1997) Biochemistry (Moscow), 62, 1191-1195.
29. Singh, R., Barden, A., Mori, T., and Beilin, L. (2001)
2. Skulachev, V. P. (1999) Biochemistry (Moscow), 64, 1418-1426. Diabetologia, 44, 129-146.
3. Longo, V. D., Mitteldorf, J., and Skulachev, V. P. (2005)
30. Lander, H. M., Tauras, J. M., Ogiste, J. S., Hori, O., Moss, R. Nat. Rev. Gen., 6, 866-872.
A., and Schmidt, A. M. (1997) J. Biol. Chem., 272, 17810-17814.
4. Skulachev, V. P. (2012) Biochemistry (Moscow), 77, 689-706.
31. Anisimov, V. N. (2003) Molecular and Physiological Mechanisms
5. Dilman, V. M. (1978) Mech. Ageing. Dev., 8, 153-173. of Aging [in Russian], 1st Edn., Nauka, St. Petersburg.
6. Dilman, V. M. (1982) Master Biological Clock [Russian
32. Anisimov, V. N., Popovich, I. G., Zabezhinski, M. A.,
Anisimov, S. V., Vesnushkin, G. M., and Vinogradova, I. A.
7. Comfort, A. (1979) The Biology of Senescence, 3rd Edn.,
(2006) Biochim. Biophys. Acta, 1757, 573-589.
33. Karasek, M. (2007) J. Physiol. Pharmacol., 58, 105-113.
8. George, J. C., Bada, J., Zeh, J., Scott, L., Brown, S. E.,
34. Pierpaoli, W., and Bulian, D. (2005) Ann. N. Y. Acad. Sci.,
O’Hara, T., and Suydam, R. (1999) Can. J. Zool., 77, 571-580. 1057, 133-144.
9. Sivan, S. S., Van El, B., Merkher, Y., Schmelzer, C. E.,
35. Skulachev, V. P., Severin, F. F., and Feniouk, B. A. (2013) Life
Zuurmond, A. M., Heinz, A., Wachtel, E., Varga, P. P.,
without Aging [in Russian], EKSMO, Moscow (in print).
Lazary, A., Brayda-Bruno, M., and Maroudas, A. (2012)
36. Severin, F. F., and Skulachev, V. P. (2009) Uspekhi Gerontol.,
Biochim. Biophys. Acta, 1820, 1671-1677. 22, 37-48.
10. Robinson, A. B., and Robinson, N. E. (1991) Proc. Natl.
37. Zhang, G., Li, J., Purkayastha, S., Tang, Y., Zhang, H., Yin,
Acad. Sci. USA, 88, 8880-8884.
Y., Li, B., Liu, G., and Cai, D. (2013) Nature, 497, 211-216.
11. Robinson, N. E., and Robinson, A. B. (2004) Mech. Ageing.
38. Namiki, M. (1988) Adv. Food Res., 32, 115-184. Dev., 125, 259-267.
39. Dworkin, J. P., and Miller, S. L. (2000) Carbohydrate Res.,
11a. Robinson, N. E., and Robinson, A. B. (2004) Molecular329, 359-365. Clocks: Deamination of Asparaginyl and Glutaminyl Residues in
40. Swamy, M. S., Tsai, C., Abraham, A., and Abraham, E. C. Peptides and Proteins, Althouse Press, Cave Junction, Oregon
(1993) Exp. Eye Res., 56, 177-185.
(http://book.deamidation.org/MolecularClocks.pdf).
41. Ledesma-Osuna, A. I., Ramos-Clamont, G., and Vazquez-
12. Robinson, N. E., and Robinson, A. B. (2001) Proc. Natl.
Moreno, L. (2008) Acta Biochim. Polon., 55, 491-497. Acad. Sci. USA, 98, 4367-4372.
42. Song, X., Bao, M. M., Li, D. D., and Li, Y. M. (1999)
13. Krautwald, M., and Munch, G. (2010) Exp. Gerontol., 45, Mech. Ageing Dev., 108, 239-251.
43. Liang, Y. X., Wang, Z., Li, D. D., Jiang, J. M., and Shao,
14. Biemel, K. M., Friedl, D. A., and Lederer, M. O. (2002) J.
R. G. (2003) Biomed. Environ. Sci., 16, 323-332. Biol. Chem., 277, 24907-24915.
44. Deng, H. B., Cheng, C. L., Cui, D. P., Li, D. D., Cui, L.,
15. Maillard, L. (1912) C. R. Hebd. Seances Acad. Sci., 154, 66-68.
and Cai, N. S. (2006) Biomed. Environ. Sci., 19, 432-438.
16. Hodge, J. E. (1953) J. Agr. Food Chem., 1, 928-943.
45. Lu, J., Wu, D. M., Zheng, Y. L., Hu, B., and Zhang, Z. F.
17. Munch, G., Schicktanz, D., Behme, A., Gerlach, M.,
(2010) Brain Pathol., 20, 598-612.
Riederer, P., Palm, D., and Schinzel, R. (1999) Nat.
46. Mao, G. X., Deng, H. B., Yuan, L. G., Li, D. D., Li, Y. Y.,
Biotechnol., 17, 1006-1010.
and Wang, Z. (2010) Biomed. Environ. Sci., 23, 161-166.
18. Monnier, V. M., Stevens, V. J., and Cerami, A. (1981) Prog.
47. Bujak-Gizycka, B., Suski, M., Olszanecki, R., Madej, J.,
Food Nutr. Sci., 5, 315-327.
and Korbut, R. (2009) Folia Medica Cracoviensia, 50, 21-33
19. Dyer, D. G., Dunn, J. A., Thorpe, S. R., Bailie, K. E.,
Lyons, T. J., McCance, D. R., and Baynes, J. W. (1993) J.
48. Anisimov, V. N. (2008) Molecular and Physiological MechanismsClin. Invest., 91, 2463-2469. of Aging [in Russian], 2nd Edn., Nauka, St. Petersburg.
20. Sell, D. R., Kleinman, N. R., and Monnier, V. M. (2000)
49. Stern, D., Yan, S. D., Yan, S. F., and Schmidt, A. M. FASEB J., 14, 145-156.
(2002) Adv. Drug. Deliv. Rev., 54, 1615-1625.
21. DeGroot, J., Verzijl, N., Wenting-Van Wijk, M. J., Bank, R.
50. Sims, G. P., Rowe, D. C., Rietdijk, S. T., Herbst, R., and
A., Lafeber, F. P., Bijlsma, J. W., and TeKoppele, J. M.
Coyle, A. J. (2010) Annu. Rev. Immunol., 28, 367-388.
(2001) Arthritis Rheum., 44, 2562-2571.
51. Nielsen, J. M., Kristiansen, S. B., Norregaard, R.,
22. Araki, N., Ueno, N., Chakrabarti, B., Morino, Y., and
Andersen, C. L., Denner, L., Nielsen, T. T., Flyvbjerg, A.,
Horiuchi, S. (1992) J. Biol. Chem., 267, 10211-10214.
and Botker, H. E. (2009) Eur. J. Heart Fail, 11, 638-647.
23. Frye, E. B., Degenhardt, T. P., Thorpe, S. R., and Baynes,
52. Kim, J., Wan, C. K., O’Carroll, S. J., Shaikh, S. B., and
J. W. (1998) J. Biol. Chem., 273, 18714-18719.
Nicholson, L. F. B. (2012) J. Neurosci. Res., 90, 1136-1147.
MATERIAL SAFETY DATA SHEET CHEMICAL PRODUCT AND COMPANY IDENTIFICATION CESI Chemical (a Flotek Industries, Inc. company) 1004 S. Plainsman Road, Marlow, OK 73055 2. COMPOSITION/INFORMATION ON INGREDIENTS Ingredient Peroral 6.48 ml/kg (rat) Percutaneous 8.0 ml/kg (rabbit) Fathead Minnow 8,300 mg/l (96 h) Daphnia 7,550 mg/l (48 h) Dermal >5g/kg (rabbit) Oral >5g/kg (rat)
 N-e-carboxy-
N-e-carboxy-