He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Neurologie.usz.ch
Copyright 2003 by the American Psychological Association, Inc.
Opposite Turning Behavior in Right-Handers and Non-Right-Handers
Suggests a Link Between Handedness and Cerebral
University Hospital Geneva and University Hospital Zurich
Department of Veterans Affairs Spark M. Matsunaga Medical
The strong right hand preference in humans remains a riddle; no lateralized behavior other than finefinger dexterity relates to it. The relation between handedness and language dominance may be farweaker than currently judged; after all, both right-handers and non-right-handers utilize the left brain forspeech. There is, however, a lateralized motor preference in animals, turning behavior, that is stronglyassociated with hemispheric dopamine (DA) asymmetries. Turning consistently occurs towards the sidewith less DA. The authors tested 69 right-handers and 24 non-right-handers with a device recordingspontaneous turning behavior for 20 hr within 3 days. Findings indicate that right-handers preferredleft-sided turning and non-right-handers preferred right-sided turning. This result suggests a link betweenhandedness and DA asymmetries.
Across cultures, humans have a strong right-hand preference,
In vertebrates, however, there exists another stable intraindi-
reportedly manifest for at least the past 5,000 years of human
vidual motor preference, turning behavior, which is strongly asso-
history (Coren & Porac, 1977). However, there seems to be no
ciated with hemispheric dopamine (DA) asymmetries. In animals,
clear phylogenetic root to this human right-bias since handed-
the direction is ipsilateral to the hemisphere with the less active
ness— or “pawedness”—appears equally distributed in most of the
DA system, or in other words, contralateral to the hemisphere with
highest primates (Hamilton & Vermeire, 1988; see Palmer, 2002
the more active DA system (Glick, 1983; Patino, Garcia-Munoz, &
for a meta-analysis). The preponderance of human right-
Freed, 1995). This relation is so well established that without
handedness has remained a riddle and is not known to be related to
quantitative DA measurements, turning behavior alone is taken as
any motor or cognitive behavior, other than measures of fine finger
proof of DA asymmetries (Brunner & Gattaz, 1995; Lindemann,
dexterity (e.g., Bryden, Pryde, & Roy, 2000; Triggs, Calvanio,
Lessenich, Ebert, & Loscher, 2001). In humans, there is one
Levine, Heaton, & Heilman, 2000). In view of the fact that both
known pathological condition, asymmetric Parkinson’s disease, in
right-handers and about 70% of non-right-handers utilize the left
which one hemisphere contains less DA than the other. Consistent
brain for speech (Knecht et al., 2000; Pujol, Deus, Losilla, &
with animal findings, such patients turn away from the hemisphere
Capdevila, 1999; Signore, Chaoui, Nosten-Bertrand, Perez-Diaz,
that is more intact (Bracha, Shults, Glick, & Kleinman, 1987).
& Marchaland, 1991), a direct link between handedness and lan-
Because one motor side preference, turning behavior, is associ-
ated with asymmetries in the DA system, it was tempting topropose that another motor side preference, handedness, mightdepend upon this system as well. This DA– handedness hypothesishas a long history in animals and, to some extent, in humans
C. Mohr, Department of Rehabilitation, University Hospital Geneva, Ge-
(Bracha, Seitz, Otemaa, & Glick, 1987; de la Fuente-Fernandez,
neva, Switzerland, and Department of Neurology, University Hospital Zurich,Zurich, Switzerland; T. Landis, Department of Neurology, University Hospital
Kishore, Calne, Ruth, & Stoessl, 2000; Fitzgerald, Ratty, Teitler,
Geneva; H. S. Bracha, National Center for PTSD, Department of Veterans
Gross, & Glick, 1993; Glick, 1983; Kooistra & Heilman, 1988;
Affairs Spark M. Matsunaga Medical and Regional Office Center, Honolulu,
Larson, Dodson, & Ward, 1989; Nielsen et al., 1997). Various
Hawaii; P. Brugger, Department of Neurology, University Hospital Zurich.
asymmetries of the volume and DA content of different parts of the
This research was supported by Grant 690610 from the Institut fu¨r
basal ganglia have been associated with handedness (Kooistra &
Grenzgebiete der Psychologie und Psychohygiene, Freiburg, Germany,
Heilman, 1988). However, a direct functional support of this
Grant 31-65096.01 from the Swiss National Science Foundation, and by
hypothesis, a relation of handedness and spontaneous turning
the Office of Research and Development, Medical Research Service,
behavior, has not been found in animals (Fitzgerald et al., 1993;
Larson et al., 1989; Nielsen et al., 1997; Westergaard & Suomi,
Correspondence concerning this article should be addressed to C. Mohr,
1996), and the only study in humans did not yield a clear-cut
Functional Brain Mapping Laboratory, University Hospital Geneva, RueMicheli-du-Crest 24, CH-1211 Geneva 14, Switzerland. E-mail: christine
picture (Bracha, Seitz, et al., 1987). These latter authors tested
hemispheric dominance, a compound measure of hand–foot– eye
preference in relation to turning behavior. They found a complex
case of removal, subjects were required to deposit the device in such a way
interaction between hemispheric dominance, turning, and gender.
as to minimize any confounding non-body-related movements. At the end
Unfortunately, the intercorrelation between hand, foot, and eye
of the 20-hr test-time, the subjects had to fill out the handedness
dominances is known to be low (Gabbard & Iteya, 1996; Graves,
1983). Thus, this study could not really answer the crucial questionof a relationship between human handedness and turning behavior.
We thus aimed to directly test human turning behavior in
We calculated a conventional laterality index score for the number of
relation to handedness in a large sample of healthy right-handers
360° turns: [(right turns minus left turns) Ϭ (right turns plus left turns)] ϫ
100 (Marshall, Caplan, & Holmes, 1975). This laterality index is a measureindependent from the individual number of turns, whereby positive values
indicate a preference to turn to the right; and negative values, to the left. The laterality index was normally distributed (Kolmogorov–Smirnov Test
d ϭ 0.05, p Ͼ .20). Gender has been found to relate differently tohemispheric dominance and turning preference (Bracha, Seitz, et al., 1987).
A total of 93 healthy subjects (48 women) with a mean age of 29.5 years
Thus, we performed a three-way analysis of variance (ANOVA) with
(range ϭ 21–59, see Table 1) were recruited by flyers and personal contact.
gender (women vs. men) and handedness groups (right-handers vs. non-
The study protocol, in accordance with the Declaration of Helsinki, was
right-handers) as between-subject variables on laterality index scores.
approved by the local human research ethics committee, and all subjects
Previous studies on language lateralization and handedness found a linear
gave signed consent before participation.
relationship between increasing left-handedness and diminished left-hemisphere dominance for language (Knecht et al., 2000); thus, we also
performed Pearson correlations between handedness raw scores and later-ality index scores. Finally, we determined individuals’ preference to turn to
Handedness was assessed with the 13-item handedness questionnaire
either the left or right side by subtracting full turns to the right from full
from Chapman and Chapman (1987) known for a high internal consistency
turns to the left. Thus, a positive value indicated a right-sided turning
and good test–retest reliability. For each item, subjects indicated whether
preference; and a negative value, a left-sided turning preference. If not
they perform the action with the right hand, left hand, or both hands. The
stated otherwise, all reported p values are two-tailed, and only significant
use of the right hand is given 1 point, of both hands 2 points, and of the left
hand 3 points. Possible scores thus range continuously from 13 (all itemsanswered with “right”) to 39 (all items answered with “left”). According tothe cut-off scores (Chapman & Chapman, 1987), the sample was subdi-
vided into right-handers (scores 13–17, n ϭ 69, 35 women) and non-right-handers (scores 18 –39, n ϭ 24, 13 women, see Table 1).
A three-way ANOVA with gender (women vs. men) and hand-
edness groups (right handers vs. non-right-handers) on age re-
We used a lightweight, rechargeable, belt-mounted device comprising a
vealed only a significant main effect for gender, F(1, 89) ϭ 4.43,
position sensor and an electronic processing circuit that monitors changes
p ϭ .04; men were older (31.1 Ϯ 7.6) than women (28.0 Ϯ 5.3; see
in the orientation of the dorsal–ventral axis (see Bracha, Seitz et al., 1987;
Bracha, Shults et al., 1987, for further technical details). Magnetic north isused as an external reference and is tracked by a compass. With this device,
partial turns (90°) in either direction are summed consecutively while theperson moves in the same direction. When four 90° quadrants have been
completed, a 360° turn is registered. If the subject suddenly moves in the
ality index revealed a significant main effect for handedness
other direction, a new measurement begins for the opposite direction. We
groups, F(1, 89) ϭ 13.30, p ϭ .0004; right-handers had a lower
assessed the number of full 360° turns to either side. Subjects were blind
laterality index than non-right-handers (Table 1). As can be seen in
to the hypothesis and specific kind of measurement. They were required towear the device during 20 hr for 3 consecutive days. At the first testing
Figure 1, right-handers turned more strongly to the left and non-
session, instructions were provided about the correct use of the device.
right-handers more strongly to the right. The difference from zero
Important instructions were that the device should be worn all day, re-
(an equal preference to turn to either side) was significant for the
moved only for sports, sleep, or activities damaging to the device. In the
right-handed group, t(68) ϭ 4.24, p Ͻ .0001 (one-tailed), and was
Table 1Demographic and Turning Measures of the Study Sample
RH ϭ right-handers; NRH ϭ non-right-handers; (L/R) ϭ number of subjects with a left-sided (L) or right-sided (R) turning preference.
Magnitude and opposite turning preferences of the two handedness groups, as shown by the mean
also significant, although less pronounced, for the non-right-
way left-handers do. If the organizational principle were guided by
handed group, t(23) ϭ 1.73, p ϭ .048 (one-tailed).
preferred hand use, our results would constitute a simple artifact. Relationship between handedness raw scores and turning index
Although future studies should address this possibility (e.g., by
limiting measurements to “controlled” environments), we do not
handedness) were positively correlated to laterality index scores,
think that it accounts for the present findings, because (a) a great
that is, more pronounced right-sided turning (r ϭ .33, p ϭ .001).
amount of data were collected in natural, outdoor environmentsand (b) the animal literature found that DA asymmetry is the final
common pathway of circling behavior (for overviews, see Glick &Ross, 1981; Pycock, 1983).
The log-linear analysis revealed a significant main effect for
The strong right-side bias of human handedness is phylogeneti-
handedness groups, 2(1, N ϭ 93) ϭ 22.72, p ϭ .000002 (more
cally unique, as animals up to the highest primates have individual
subjects were right-handed than non-right-handed) and preferred
paw preferences, but these appear equally distributed on the pop-
turning side, 2(1, N ϭ 93) ϭ 5.75, p ϭ .02 (more subjects
ulation level as to the side used (Betancur, Neveu, & Le Moal,
preferred left turns to right turns), as well as a significant interac-
1991; Hamilton & Vermeire, 1988; Palmer, 2002; Signore et al.,
tion between handedness groups and preferred turning side, 2(1,
1991). Animal studies will thus not necessarily answer questions
N ϭ 93) ϭ 15.02, p ϭ .0001 (right-handed subjects preferred left
about human handedness. While turning behavior in animals is
turns and non-right-handed subjects preferred right turns; Table 1).
widely accepted to depend on an asymmetric DA system, theneurochemical and/or neuroanatomical basis of paw preference
has received little attention. It was tempting to relate side of
The results of the present study show a reliable categorization of
turning, and implicitly DA asymmetries, to pawedness. As early as
groups of people with opposite handedness according to another
1974, Glick and Jerussi studied spatial lever press preferences in
dichotomous behavioral measure, spontaneous turning behavior
rats as a function of pawedness. These authors found spatial
toward the side opposite to the preferred hand. One might argue
lever-press preference to be related to turning side. Unfortunately,
that this preference is simply an artifact of an overactive dominant
the authors did not report preferred pawedness and turning pref-
hand (or foot). But if increased unilateral limb motor activity is
erence. In subsequent studies by the Glick group, a direct link and
associated with increased contralateral turning behavior, patients
thus support for this DA– handedness hypothesis could not be
with asymmetrical Parkinson’s disease should turn toward the
established in animals (Fitzgerald et al., 1993; Larson et al., 1989;
more rigid, symptomatic body side, which is opposite to what is
Nielsen et al., 1997; see also Westergaard & Suomi, 1996). Some
actually found (Bracha, Shults, et al., 1987). Moreover, a study
relation between these variables does exist, however, as these
with healthy right-handers showed that they spontaneously made
studies showed that, independent of side, the stronger the paw
significantly more movements with the left than the right arm
preference, the stronger the turning behavior (Nielsen et al., 1997).
(Eaton, Rothman, McKeen, & Campbell, 1998). One more alter-
Note that such a relationship was not found in our data (correlation
native might be considered, that is, the possibilities that right-
between absolute turning index score and handedness raw scores:
handers organize their personal environments differently from the
r ϭ Ϫ.02, p ϭ .84).
Moreover, DA injections in the caudate in rats produced an
Betancur, C., Neveu, P. J., & Le Moal, M. (1991). Strain and sex differ-
increased use of the preferred paw when ipsilaterally injected, but
ences in the degree of paw preference in mice. Behavioural Brain
a decreased use when contralaterally injected (Evenden & Rob-
bins, 1984). Another way of approaching this question was used in
Bracha, H. S., Seitz, D. J., Otemaa, J., & Glick, S. D. (1987). Rotational
unilateral striatal lesion studies and in studies measuring DA in
movement (circling) in normal humans: Sex difference and relationshipto hand, foot and eye preference. Brain Research, 411, 231–235.
different parts of the basal ganglia: The striatum has been found to
Bracha, H. S., Shults, C., Glick, S. D., & Kleinman, J. E. (1987). Sponta-
play an important role in contralateral paw-reaching skills and
neous asymmetric circling behavior in hemi-parkinsonism: A human
ipsilateral turning behavior (Barne´oud et al., 1995; Henderson et
equivalent of the lesioned-circling rodent behavior. Life Sciences, 40,
al., 1999), and paw preference has been related to an ipsilateral
hemispheric dominance for dopamine in the nucleus accumbens in
Brunner, J., & Gattaz, W. F. (1995). Intracerebral injection of phospho-
lipase A2 inhibits dopamine-mediated behavior in rats: Possible impli-
In humans, similar attempts to relate size and/or DA asymme-
cations for schizophrenia. European Archives of Psychiatry and Clinical
tries of basal ganglia structures to handedness have been under-
taken. The globus pallidus especially appears to be larger and to
Bryden, P. J., Pryde, K. M., & Roy, E. A. (2000). A performance measure
contain higher amounts of DA in the left hemisphere (Glick, Ross,
of the degree of hand preference. Brain and Cognition, 44, 402– 414.
& Hough, 1982; Kooistra & Heilman, 1988). Kooistra and
Cabib, S., D’Amato, F. R., Neveu, P. J., Deleplanque, B., Le Moal, M., &
Heilman, for example, analyzed the postmortem brains of 18
Puglisi-Allegra, S. (1995). Paw preference and brain dopamine asym-metries. Neuroscience, 64, 427– 432.
individuals who were healthy and found significantly larger left
Chapman, L. J., & Chapman, J. P. (1987). The measurement of handed-
globus pallidus size. Assuming right-handedness in these individ-
ness. Brain and Cognition, 6, 175–183.
uals, they speculated about a causal link between DA asymmetries
Coren, S., & Porac, C. (1977, November 11). Fifty centuries of right-
and turning behavior on the one hand and the development of limb
handedness: The historical record. Science, 198, 631– 632.
motor asymmetries on the other hand. However, according to this
de la Fuente-Fernandez, R., Kishore, A., Calne, D. B., Ruth, T. J., &
line of reasoning, one would expect right-handers to have a right-
Stoessl, A. J. (2000). Nigrostriatal dopamine system and motor lateral-
sided, but not left-sided, turning preference.
ization. Behavioural Brain Research, 112, 63– 68.
A recent study measured the unimanual and bimanual motor
Eaton, W. O., Rothman, D. B., McKeen, N. A., & Campbell, D. W. (1998).
performance of 20 right-handed healthy subjects, and, indepen-
Something sinistral going on? Asymmetry in arm movement frequency.
dently, their at-rest (18F) Fluorodopa PET uptake (de la Fuente-
Fernandez et al., 2000). The authors calculated left–right uptake
Evenden, J. L., & Robbins, T. W. (1984). Effects of unilateral
6-hydroxydopamine lesions of the caudate-putamen on skilled forepaw
asymmetries in the basal ganglia and statistically correlated them
use in the rat. Behavioural Brain Research, 14, 61– 68.
to the individual’s motor proficiency. They found right-hand per-
Fitzgerald, L. W., Ratty, A. K., Teitler, M., Gross, K. W., & Glick, S. D.
formance to correlate with left putamen dominance, but unexpect-
(1993). Specificity of behavioral and neurochemical dysfunction in the
edly bimanual performance to correlate with right caudate domi-
chakragati mouse: A novel genetic model of a movement disorder. Brain
nance. Because they did not test non-right-handers, the question of
handedness could not be addressed with this study. However, their
Gabbard, C., & Iteya, M. (1996). Foot laterality in children, adolescents,
correlation of good bimanual performance in right-handers with a
and adults. Laterality, 1, 199 –205.
relative “dominance” of the right caudate is very interesting.
Glick, S. D. (1983). Cerebral lateralization in the rat and tentative extrap-
Bimanual actions, by their inherent demand for interhemispheric
olations to man. In M. S. Myslobodsky (Ed.), Hemisyndromes: Psycho-
connectivity, are closer physiologically to axial whole-body move-
biology, neurology, psychiatry (pp. 7–23). New York: Academic Press.
ments—such as turning behavior—than are the fine unilateral
Glick, S. D., & Jerussi, T. P. (1974). Spatial and paw preferences in rats:
finger movements that usually determine handedness. If one ac-
Their relationship to rate-dependent effects of d-amphetamine. Journalof Pharmacology and Experimental Therapeutics, 188, 714 –725.
cepts a link between bimanual action and axial motion, this study,
Glick, S. D., & Ross, D. A. (1981). Lateralization of function in the rat
because of the right-sided DA dominance, might better explain our
brain basic mechanisms may be operative in humans. Trends in Neuro-
left-sided turning behavior in right-handers.
In conclusion, we found, in a large sample of right-handers and
Glick, S. D., Ross, D. A., & Hough, L. B. (1982). Lateral asymmetry of
non-right-handers during a 20-hr spontaneous turning behavior
neurotransmitters in human brain. Brain Research, 234, 53– 63.
study, that right-handers turn left and non-right-handers turn right.
Graves, R. (1983). Mouth asymmetry, dichotic ear advantage and tachis-
This finding, although indirect, constitutes the first experimental
toscopic visual field advantage as measures of language lateralization.
evidence of a link between human handedness and another dichot-
Neuropsychologia, 21, 641– 649.
omous behavioral variable, other than fine finger dexterity. We
Hamilton, C. R., & Vermeire, B. A. (1988, December 23). Complementary
also found that, primarily on the basis of animal studies, this
hemispheric specialization in monkeys. Science, 242, 1691–1694.
turning behavior is likely to be due to DA asymmetries and may
Henderson, J. M., Annett, L. E., Ryan, L. J., Chiang, W., Hidaka, S.,
thus provide a link between the DA system and the development of
Torres, E. M., & Dunnett, S. B. (1999). Subthalamic nucleus lesionsinduce deficits as well as benefits in the hemiparkinsonian rat. EuropeanJournal of Neuroscience, 11, 2749 –2757.
Knecht, S., Dra¨ger, B., Deppe, M., Bobe, L., Lohmann, H., Flo¨el, A.,
Ringelstein, E.-B., & Henningsen, H. (2000). Handedness and hemi-
Barne´oud, P., Parmentier, S., Mazadier, M., Miquet, J. M., Boireau, A.,
spheric language dominance in healthy humans. Brain, 123, 2512–2518.
Dubedat, P., & Blanchard, J. C. (1995). Effects of complete and partial
Kooistra, C. A., & Heilman, K. M. (1988). Motor dominance and lateral
lesions of the dopaminergic mesotelencephalic system on skilled fore-
asymmetry of the globus pallidus. Neurology, 38, 388 –390.
limb use in the rat. Neuroscience, 67, 837– 848.
Larson, C. F., Dodson, D. L., & Ward, J. P. (1989). Hand preferences and
whole-body turning biases of lesser bushbabies (Galago senegalensis).
lateralization of language in normal left-handed people studied by func-
Brain, Behavior and Evolution, 33, 261–267.
tional MRI. Neurology, 52, 1038 –1043.
Lindemann, S., Lessenich, A., Ebert, U., & Loscher, W. (2001). Sponta-
Pycock, C. J. (1983). Experimental model of hemi-Parkinsonism. In M. S.
neous paroxysmal circling behavior in the CI2 rat mutant: Epilepsy with
Myslobodsky (Ed.), Hemisyndromes: Psychobiology, neurology, psychi-
rotational seizures or hyperkinetic movement disorder? Experimentalatry (pp. 69 –90). New York: Academic Press.
Signore, P., Chaoui, M., Nosten-Bertrand, M., Perez-Diaz, F., & Marcha-
Marshall, J., Caplan, D., & Holmes, J. (1975). The measure of laterality.
land, C. (1991). Handedness in mice: Comparison across eleven inbred
Neuropsychologia, 13, 315–321.
strains. Behavior Genetics, 21, 421– 429.
Nielsen, D. M., Visker, K. E., Cunningham, M. J., Keller, R. W., Jr, Glick,
Triggs, W. J., Calvanio, R., Levine, M., Heaton, R. K., & Heilman, K. M.
S. D., & Carlson, J. N. (1997). Paw preference, rotation, and dopamine
(2000). Predicting hand preference with performance on motor tasks.
function in Collins HI and LO mouse strains. Physiology & Behavior,
Westergaard, G. C., & Suomi, S. J. (1996). Hand preference for a bimanual
Palmer, A. R. (2002). Chimpanzee right-handedness reconsidered: Evalu-
task in tufted capuchins (Cebus apella) and rhesus macaques (Macaca
ating the evidence with funnel plots. American Journal of Physicalmulatta). Journal of Comparative Psychology, 110, 406 – 411. Anthropology, 118, 191–199.
Patino, P., Garcia-Munoz, M., & Freed, C. R. (1995). Electrophysiology of
ventromedial striatal neurons during movement. Brain Research Bulle-
Pujol, J., Deus, J., Losilla, J. M., & Capdevila, A. (1999). Cerebral
Johannes Kepler Univ. of Linz & Inst. of Problems of Chemical Physics of Russian: Fabrication of “green” organic field-effect transistors N. Marjanovic M. Irimia-Vladu and colleagues from the Johannes Kepler University of Linz & Institute of Problems of Chemical Physics of Russian present this work in the frame of the “green potentiality” of organic electr
FIGURE 3-24 Approach to hyperkalemia: hyperkalemiawith reduced glomerular filtration rate(GFR). Normokalemia can be maintained in patients who consume normal quantitiesof potassium until GFR decreases to lessthan 10 mL/min; however, diminished GFRpredisposes patients to hyperkalemia fromexcessive exogenous or endogenous potassi-um loads. Hidden sources of endogenous andexogenous potassi
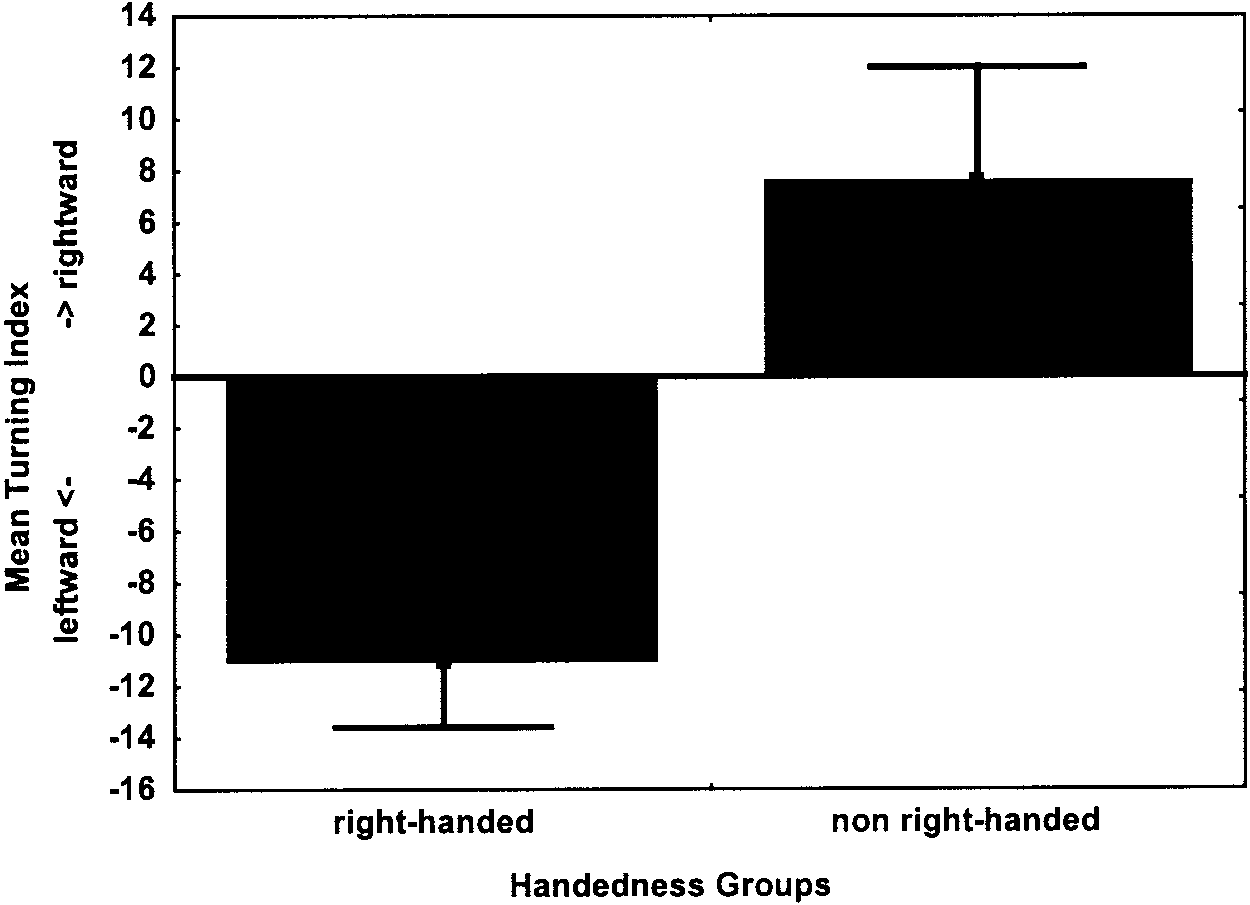 Magnitude and opposite turning preferences of the two handedness groups, as shown by the mean
also significant, although less pronounced, for the non-right-
way left-handers do. If the organizational principle were guided by
handed group, t(23) ϭ 1.73, p ϭ .048 (one-tailed).
Magnitude and opposite turning preferences of the two handedness groups, as shown by the mean
also significant, although less pronounced, for the non-right-
way left-handers do. If the organizational principle were guided by
handed group, t(23) ϭ 1.73, p ϭ .048 (one-tailed). whole-body turning biases of lesser bushbabies (Galago senegalensis).
whole-body turning biases of lesser bushbabies (Galago senegalensis).