He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Evol_56_1018.1976_1981.tp
Evolution, 56(10), 2002, pp. 1976–1981
WOLBACHIA INTERACTIONS THAT DETERMINE
Department of Ecology and Evolutionary Biology, Brown University, Box G-W, Providence, Rhode Island 02912
1E-mail: Adam[email protected]
We have recently described a mutualistic symbiosis in which Wolbachia bacteria were shown to improve
the fitness of some Drosophila melanogaster stocks. Wolbachia did not extend longevity in all Drosophila genotypes,even though 16s rDNA sequences indicated that our Drosophila stocks were infected with the same Wolbachia strain. Here, we use reciprocal hybrid crosses between two Drosophila strains, one that lived longer with Wolbachia (Z53)and one that did not (Z2), to investigate the inheritance of the survival phenotype and its dependence on the hostgenotype, sex, and mating conditions. Wolbachia’s positive effects were more apparent in hybrid flies than in parentalflies, ruling out exclusive maternal inheritance or the dependence of the survival phenotype on Wolbachia straindifferences. The Wolbachia survival effects were more apparent in single-sex cages, where courtship and mating werenot permitted. In these cages, nearly all flies with Wolbachia lived longer than uninfected flies, even though strainZ2 showed no Wolbachia effect in mixed-sex mating cages. We used comparisons between single- and mixed-sexcages to estimate the cost of reproduction for both sexes. Our data suggest that Wolbachia infection may increase theinferred cost of reproduction, particularly in males. Wolbachia can even produce a positive survival effect almost aslarge as the negative survival effect associated with reproduction. We discuss the implications of our experiments forthe study of insect symbioses.
Cost of reproduction, hybrid, mortality, survival, symbiosis.
␣-proteobacteria Wolbachia and Drosophila is in transition from parasitic to
thought to infect millions of insect species (Werren et al.
mutualistic (A. J. Fry, M. R. Palmer, and D. M. Rand, unpubl.
1995a; Jeyaprakash and Hoy 2000; Werren and Windsor
ms.). To better understand these survival effects, we used
2000; Jiggins et al. 2001a). Their widespread distribution
reciprocal hybrid crosses between two Drosophila strains,
among insects makes them one of the most common infec-
one that lived longer with Wolbachia (Z53) and one strain
tious microorganisms. These bacteria inhabit the reproductive
(Z2) that did not show a significant Wolbachia effect. This
tissues of their hosts (Dobson et al. 1999), where they induce
design allowed us to compare the survival effects of the Wol-
a number of reproductive modifications intended to enhance
bachia strains found in Z53 and Z2 as each hybrid had a
their transmission through females. Their effects include the
different Wolbachia strain. We also investigated Wolbachia’s
induction of parthenogenesis (Stouthamer et al. 1993), fem-
effects in both inbred (Z53 and Z2) and outbred (hybrid)
inization of genetic males (Rousset et al. 1992), male-killing
genotypes and we manipulated the mating conditions to un-
(Jiggins et al. 1998; Hurst et al. 1999), and cytoplasmic in-
derstand Wolbachia’s interaction with reproduction. This was
compatibility (CI; Caspari and Watson 1959; Yen and Barr
done because courtship and mating are known to affect Dro-
1971; Fine 1978; Hoffmann et al. 1986, 1990). Wolbachiasophila survival (e.g., Fowler and Partridge 1989; Partridge
does not induce strong reproductive modifications in Dro-
and Fowler 1990; Chapman et al. 1995) and could interact
sophila melanogaster, and this has led to the suggestion that
with Wolbachia’s effects on survival. Wolbachia may benefit D. melanogaster fitness (Hoffmann etal. 1994, 1998; Solignac et al. 1994). Attempts to identify
these benefits revealed that, in the field, Wolbachia couldimprove survival, but the positive survival effects depended
on the population background and the location of the field
The Z53 and Z2 D. melanogaster stocks were originally
site (Olsen et al. 2001). In the laboratory, we found that
collected in Zimbabwe, Africa. These laboratory strains were
Wolbachia could significantly improve both survival and fe-
started as isofemale lines, have been in laboratory culture for
male fecundity, and these effects depended on the host ge-
several hundred generations, and have been maintained at
notype (A. J. Fry, M. R. Palmer, and D. M. Rand, unpubl.
moderate population size (100–200 pairs) in the laboratory
ms.). DNA sequences from 16s rDNA (O’Neill et al. 1992)
of D. M. Rand. They also carry the endosymbiotic bacterium
and the ftsZ cell cycle gene (Holden et al. 1993; Werren et
W. pipientis. Marc Tatar (Brown University) provided Ri-
al. 1995b) suggested that our Wolbachia strains were iden-
RedE, an uninfected control strain. We created paired Wol-
tical. However, recent reports of recombination between dif-
bachia-infected (W) and tetracycline-treated (T) experimen-
ferent Wolbachia strains preclude exact strain identification
tal lines with a standard protocol (Hoffmann et al. 1994;
unless a large number of relevant loci are compared (Werren
Poinsot and Mercot 1997). The uninfected fly lines are called
et al. 1995b; Werren and Bartos 2001; Jiggins et al. 2001b).
(T) because they were treated with the antibiotic tetracycline
The positive fitness effects we found could have important
for two generations to remove Wolbachia (0.25 mg/ml tet-
implications for the maintenance of Wolbachia infection in
racycline in water added to Carolina dry food in a 1:1 mix). D. melanogaster and may indicate that the symbiosis between
The Wolbachia-infected (W) lines received identical food and
᭧ 2002 The Society for the Study of Evolution. All rights reserved.
TABLE 1. Proportional hazards analysis of females from fly strains
TABLE 2. Risk ratios of Wolbachia-infected (W) to tetracycline-treat-
Z53, Z2, and their hybrids. Mating environments were mixed- or sin-
ed (T ) lines from two different mating environments, mixed- and sin-
gle-sex. Wolbachia refers to Wolbachia-infected or tetracycline-treated
gle-sex, left columns. Ratios greater than one indicate W lived longer
flies. Likelihood-ratio (LR) chi-squared values and associated proba-
than T and measure Wolbachia’s survival benefits. The two right col-
umns are risk ratios of survival in single-sex to mixed-sex cages fordifferent infection states (T or W) and are a rough measure of the
benefits to survival obtained by not reproducing. W and T or mixed-and single-sex cages were compared with log-rank tests and signifi-
cance (P Ͻ 0.001) is indicated by an asterisk.
environment, except tetracycline was not added. The infec-
tion status of all lines was confirmed with Wolbachia-specific
16S rRNA polymerase chain reaction primers (O’Neill et al.
1992). To insure that differences between T and W lines were
not due to tetracycline treatment, we used two controls. First,
we treated an uninfected fly strain, RiRedE, with tetracyclineto see if treatment affected survival. Second, we held ex-perimental flies for two generations on standard corn-meal
together in one cage. Single-sex environments were con-
laboratory food after treatment, but before the experimental
structed with 100 virgin flies of only one sex per cage. There
crosses, to minimize maternal effects of treatment.
were three replicate cages for each cross, sex, and mating
Twenty-five pairs of parental flies from the T and W lines
environment combination. Our treatment control, RiRedE,
were placed into food bottles and held until approximately
was reared in the mixed-sex environment only.
120–150 eggs were deposited. We controlled egg density
In total, we monitored 78 demography cages and scored
because W females from Z53 can lay more eggs than T fe-
the survival of more than 10,000 flies. Statistical analyses
males (A. J. Fry, M. R. Palmer, and D. M. Rand, unpubl.
were performed using the JMP statistical package (SAS In-
ms.), which could influence larval density and in turn affect
stitute 1995) and a semiparametric proportional hazards sta-
Drosophila development time (Gonzalez-Candelas et al.
tistical model (Cox 1972). In our model, the dependent var-
1990), survival (Buck et al. 1993), and Wolbachia infection
iable was time of death measured to the nearest 48 h, with
levels (Hoffmann et al. 1998). Eggs were allowed to develop
host genotype (four crosses), infection status (T or W), mat-
and virgin adults collected and paired into one of four cross
ing environment (mixed- or single-sex), and their interactions
types: two hybrid crosses, Z53 male ϫ Z2 female and Z2
male ϫ Z53 female, and two parental crosses, Z53 ϫ Z53and Z2 ϫ Z2. To avoid confounding effects of cytoplasmic
incompatibility on survival, all crosses were made with flies
The survival of female flies from Z53, Z2, and their hybrids
of the same infection status; that is, we did not cross T and
depended on interactions between Wolbachia’s effects on sur-
W flies. Twenty-five pairs of adults from each cross were
vival, the host genotype, and whether courtship and mating
placed into food bottles until 120–150 eggs were deposited.
occurred (Table 1). A proportional hazards analysis of male
Virgin F1 adults from these crosses were collected over a
survival was nearly identical except the three-way interaction
24-h period and used to initiate demography cages.
was not significant (likelihood-ratio 2 ϭ 5.82, df ϭ 3, P ϭ0.12). Due to the large number of pairwise comparisons be-
tween T and W flies, we show only some of the more im-
The demography cages were constructed from quart-serv-
portant comparisons. The complete dataset is summarized in
ing plastic containers with a screened lid; a side coupling the
same dimension as a standard food vial; and a double-walled,
The survival curves from the four crosses were signifi-
rubber side entrance, made from bicycle inner tube. The de-
cantly heterogeneous for females and males (Fig. 1), regard-
mography cages were kept in a walk-in incubator on a 12L:
less of Wolbachia infection status, in both mixed- and single-
12D photoperiod at 25ЊC and 40% relative humidity. Fresh
sex cages. Hybrid flies survived longer than flies from either
food vials we added to the cages every other day, when dead
parental strain (Fig. 1). Wolbachia had a pronounced survival
flies were removed with an aspirator, sexed, and counted.
effect in hybrid flies and flies reared in the single-sex cages
Because courtship and mating can affect Drosophila survival
(Table 2). We found that Wolbachia affected the survival of
and could interact with Wolbachia’s effects on survival
hybrid flies more than parental flies. Hybrids that carry Wol-
(Fowler and Partridge 1989; Partridge and Fowler 1990;
bachia survived significantly longer than T hybrids, even
Chapman et al. 1995), we scored survival in two different
though all hybrids had at least one parent (Z2) that did not
mating environments. Mixed-sex environments were con-
show a significant Wolbachia survival effect (Fig. 2). The
structed with 100 virgin F1 males and 100 virgin females
exception involves male hybrids with a Z2 father. Here, the
Survival curves from females (A) and males (B) from
parental crosses and hybrids. Data are from Wolbachia-infected (W)flies reared in mixed-sex mating cages. Symbols: Z53 ϫ Z53, filledtriangle; Z53 ϫ Z2, square; Z2 ϫ Z53, inverted triangle; Z2 ϫ Z2,diamond. (A) Females are significantly heterogeneous by log-rank(LR) test (2 ϭ 254, df ϭ 3, P Ͻ 10Ϫ3), (B) as are males (LR 2
ϭ 270, df ϭ 3, P Ͻ 10Ϫ3).
survival curves for T and W flies were not different in mixed-sex cages (Table 2). However, in single-sex cages, W flieslived longer than T flies, even if W and T flies were notdifferent in the mixed-sex cages as was the case for Z53males and both sexes of Z2 (Fig. 3). There were no significant
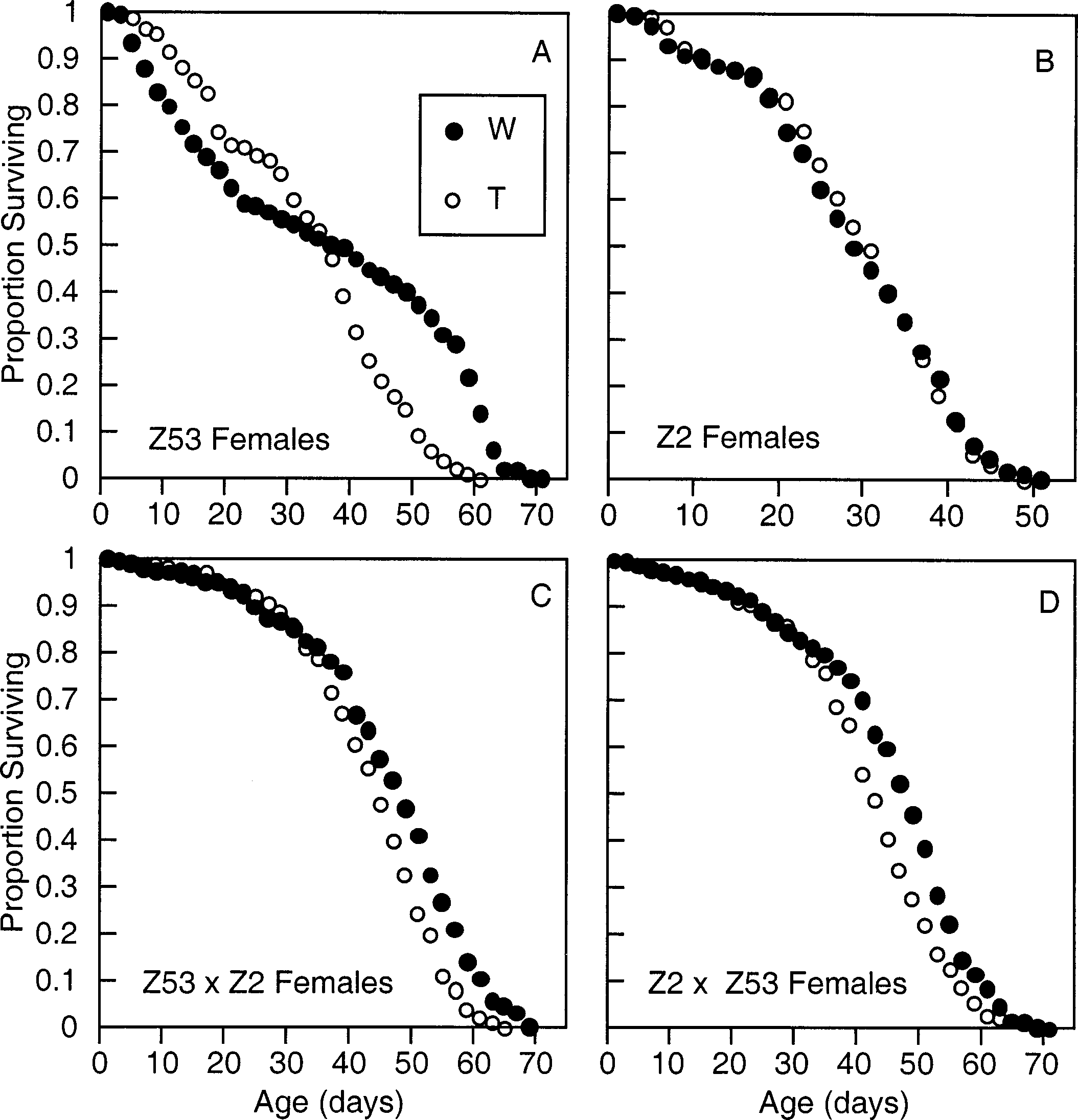
Comparison of survival curves from Wolbachia-infected
survival effects associated with tetracycline (T) treatment in
(W, closed circles) and tetracycline-treated (T, open circles) flies
our treatment control, RiRedE (P Ͼ 0.05 for both sexes by
from four genotypes. Data are from females reared in mixed-sex
log-rank test), indicating that tetracycline treatment was not
cages. (A) LR 2 ϭ 52.28, df ϭ 1, P Ͻ 10Ϫ3; (B) LR 2 ϭ 0.05,
directly responsible for the survival effects we observed. P ϭ 0.89; (C) LR 2 ϭ 69.96, P Ͻ 10Ϫ3; (D) LR 2 ϭ 15.59, P Ͻ10Ϫ3. Drosophila survival depended on strong interactions be-
tween Wolbachia infection, host genotype, and the matingconditions experienced by both male and female flies (Table1). The parental strains Z53 and Z2 have been in laboratoryculture for several hundred generations, they are inbred andhave probably accumulated mutations affecting survival. Notsurprisingly, hybrid flies were longer-lived than their parentsin both mixed- (Fig. 1) and single-sex mating environments. With one exception, the hybrids also showed a significantpositive Wolbachia effect, even though each hybrid had atleast one parent (Z2) that did not. The presence of a Wol-bachia effect in nearly all hybrids suggests that there maybe a relationship between inbreeding and the expression ofthe Wolbachia survival effect. Other Wolbachia phenotypiceffects such as the expression of cytoplasmic incompatibilityand the fidelity of maternal transmission are more pronouncedin laboratory stocks than in field populations (Hoffmann etal. 1990; Turelli and Hoffmann 1995). Drosophila fitness canbe adversely affected by inbreeding (e.g., Miller et al. 1993;Fry et al. 1998; Aspi 2000). As Wolbachia infection spreadsthroughout a host population, genetic variation at other cy-toplasmically inherited molecular markers can be reduced,such as mitochondrial DNA variation in D. simulans (e.g.,Ballard 2000). Thus, the interaction between Wolbachia andinbreeding could be important for predicting the equilibrium
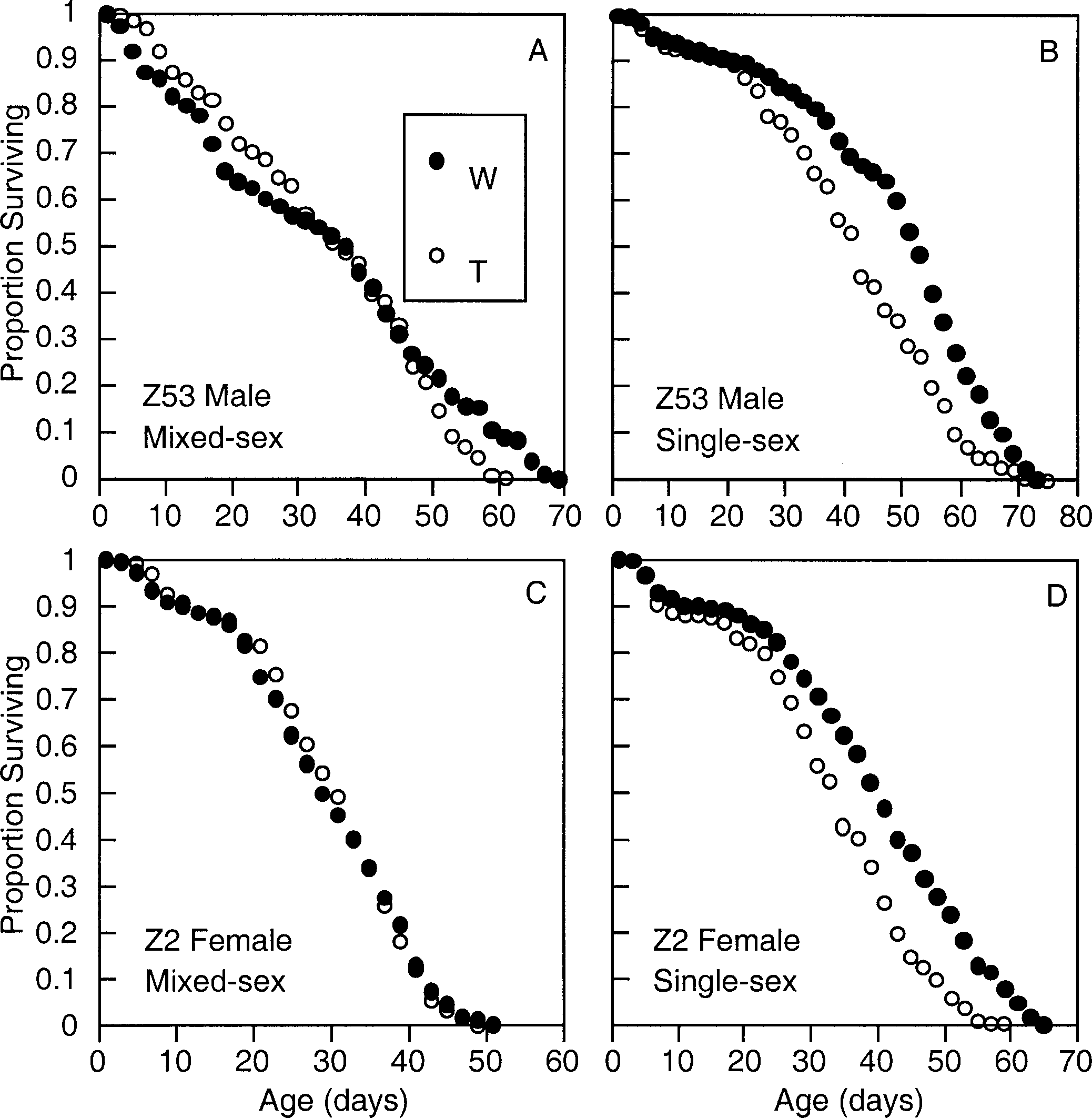
Effects of Wolbachia in mixed- and single-sex mating cag-
es. Comparisons between Wolbachia-infected (W, closed circles)
frequency and spread of Wolbachia infection in natural pop-
and tetracycline-treated (T, open circles) flies. (A) LR 2 ϭ 3.61,
df ϭ 1, P ϭ 0.05; (B) LR 2 ϭ 35.40, df ϭ 1, P Ͻ 10Ϫ3; (C) LR
There is little evidence to suggest that the positive survival
2 ϭ 0.05, df ϭ 1, P ϭ 0.89; (D) LR 2 ϭ 48.83, df ϭ 1, P Ͻ
effects associated with Wolbachia are uniparentally inherited.
TABLE 3. Mortality model analysis from Z53, Z2, and their hybrids
TABLE 4. Mortality model analysis from Z53, Z2, and their hybrids
from the mixed-sex mating cages. See Discussion for explanation of
from the single-sex mating cages. See Discussion for explanation of
models and symbols. Values give the proportional contribution of that
models and symbols. Values give the proportional contribution of that
parameter to the total difference in mortality between tetracycline-
parameter to the total difference in mortality between tetracycline-
treated (T ) and Wolbachia-infected (W) flies.
treated (T ) and Wolbachia-infected (W) flies.
However, more crosses will be needed to determine the exactinheritance pattern. We reported previously that 16S rRNAsequences from our Wolbachia strains were identical, whichimplicated the host genome in the differential expression of
that Wolbachia can decrease sperm production in D. simulans.
these Wolbachia survival effects (A. J. Fry, M. R. Palmer,
It is unclear whether a similar mechanism increases the cost
and D. M. Rand, unpubl. ms.). However, we note that recent
of reproduction in our W males. Our data reject the notion
reports of recombination between some Wolbachia strains
that Wolbachia’s positive survival effects are achieved by
(Jiggins et al. 2001b; Werren and Bartos 2001) make it nec-
decreasing the cost of reproduction. This is consistent with
essary to obtain sequences from a large number of Wolbachia
our earlier result that found Wolbachia could improve sur-
loci to determine if the strains are identical by descent. With
vival without decreasing female fecundity. In fact, Wolbachia
complete Wolbachia genomes currently being sequenced
improved both survival and fecundity of female flies (A. J.
(e.g., Slatko et al. 1999), it should soon be possible to de-
Fry, M. R. Palmer, and D. M. Rand, unpubl. ms.). The final
termine the extent to which different Wolbachia strains and
point to make about Table 2 is that by comparing Wolbachia’s
host genomes interact to produce the various Wolbachia phe-
effects on survival with the inferred cost of reproduction, we
find that Wolbachia’s positive effects can be substantial. For
The Wolbachia survival effects in our experiment showed
example, the positive effect of Wolbachia (25%) is close to
a strong dependence on the mating cage. W flies reared in
half the inferred cost of reproduction (58%) in Z53 females
single-sex cages lived longer than T flies, even if T and W
(Table 2). We suggest that Wolbachia effects of this mag-
flies were the same in the mixed-sex cages. The benefits of
nitude are probably important determinants of life-history
Wolbachia in the two environments can be compared using
evolution in chronically infected host populations. Whether
risk ratios (Table 2). For example, neither females nor males
these positive and negative survival effects might be offset-
from Z2 show a significant Wolbachia effect in the mixed-
ting in natural populations is uncertain and requires addi-
sex environment (1.01 and 1.06 risk ratios, respectively). In
the single-sex environment, however, there is a 21% and 19%
To understand how Wolbachia affects survival, we used
benefit to Wolbachia infection. Aside from Z2 and Z53 males,
maximum-likelihood analyses (Pletcher 1999) to compare es-
however, the effect of Wolbachia on survival is about the
timated mortality models from W and T flies (Tables 3, 4).
same in the mixed- and single-sex cages. For example, in
In most cases, similar models described T and W mortality
Z53 females, Wolbachia’s positive effect is about 25% under
curves from the same genotype and sex. The inferred models
were either two-parameter Gompertz models, which describe
The right two columns of Table 2 compare fly survival in
a simple exponential increase in mortality rate with age, or
mixed- and single-sex cages. If we consider these risk ratios
Gompertz-Makeham models (Vaupel and Yashin 1985) that
as a very rough index of the cost of reproduction, we can
include an additional parameter (C) to describe age-indepen-
compare Wolbachia’s effects on fly survival with the effects
dent mortality. The proportional contribution of each param-
of courtship, mating, and egg laying on fly survival. Inspec-
eter to the total difference in mortality was determined using
tion of these ratios indicates several things. First, nearly all
the method of Pletcher et al. (2000). The results are shown
of the flies experience a significant decrease in survival when
in Table 3 for mixed-sex mating cages and Table 4 for single-
courtship and mating occur. This has been well documented
sex cages. Our results indicate that Wolbachia infection can
in Drosophila and presumably represents a physiological cost
contribute to age-independent mortality. For example, 22.6%
to reproduction (e.g., Fowler and Partridge 1989; Partridge
of the total difference in mortality between T and W flies for
and Fowler 1990; Chapman et al. 1995). Second, there is a
Z53 females is due to an age-independent (C) mortality con-
trend suggesting that Wolbachia infections can contribute to
tribution. More commonly, though, Wolbachia affected both
the cost of reproduction. This cost appears much greater for
the rate of aging (slope, ), and the initial mortality rate
the parental Z53 and Z2 males than for any female. Again,
(intercept, ␣). Our data suggest that Wolbachia can affect the
this effect could be related to the level of inbreeding in pa-
mortality schedules of flies in ways that are difficult to pre-
rental males because hybrid males show no cost of repro-
dict. Clearly, Wolbachia but must be controlled in experi-
duction, infected or not (Table 2). Snook et al. (2000) showed
mental investigations of arthropod fitness.
males is mediated by male accessory gland products Nature 373:241–244.
We found strong evidence that survival in D. melanogaster
Cox, D. R. 1972. Regression models and life tables J. R. Stat. Soc.
depends on interactions between Wolbachia infection, host
genotype, host sex, and whether reproduction occurs. Wol-
Dobson, S. L., K. Bourtzis, H. R. Braig, B. F. Jones, W. Zhou, F.
Rousset, and S. L. O’Neill. 1999. Wolbachia infections are dis-
bachia had a significant positive influence on survival, and
tributed throughout insect somatic and germ line tissues. Insect
treatment to remove Wolbachia decreased survival by as
much as 25% in mixed-sex cages where mating occurred. In
Fine, P. E. M. 1978. On the dynamics of symbiote-dependent cy-
the single-sex cages where mating was not permitted, Wol-
toplasmic incompatibility in Culicine mosquitoes. J. Invertebr. Pathol. 30:10–18. bachia’s effects on survival were even more pronounced and
Fowler, K., and L. Partridge. 1989. A cost of mating in female fruit
increased survival in all flies assayed. In addition to mating
cage, the host genotype also determined whether Wolbachia
Fry, J. D., S. L. Heinsohn, and T. F. Mackay. 1998. Heterosis for
affected survival. W hybrid flies survived longer than T flies,
viability, fecundity, and male fertility in Drosophila melano-
even though all hybrids had at least one parent that did not
gaster: comparison of mutational and standing variation. Ge-netics 148:1171–1188.
show a Wolbachia survival effect. Our data suggest there may
Gonzalez-Candelas, F., J. L. Mensua, and A. Moya. 1990. Larval
be a relationship between the level of inbreeding in the host
competition in Drosophila melanogaster: effects on development
and the expression of Wolbachia-induced phenotypes. A com-
parison of Wolbachia’s effects on survival with the inferred
Hoffmann, A. A., M. Turelli, and G. M. Simmons. 1986. Unidi-
rectional incompatibility between populations of Drosophila si-
costs of reproduction in our flies revealed that Wolbachia’s
mulans. Evolution 40:692–701.
positive effect on survival can be almost as large as the neg-
Hoffmann, A. A., M. Turelli, and L. G. Harshman. 1990. Factors
ative effect of reproduction. There was also a trend suggesting
affecting the distribution of cytoplasmic incompatibility in Dro-
that Wolbachia infection contributes to the cost of reproduc-
sophila simulans. Genetics 126:933–948.
tion in both sexes, although this effect appears greater in
Hoffmann, A. A., D. J. Clancy, and E. Merton. 1994. Cytoplasmic
incompatibility in Australian populations of Drosophila melan-
males than in females, and a cost is not observed in hybrid
ogaster. Genetics 136:993–999.
males. Because Wolbachia can have both positive and neg-
Hoffmann, A. A., M. Hercus, and H. Dagher. 1998. Population
ative effects on host survival (e.g., Min and Benzer 1997;
dynamics of the Wolbachia infection causing cytoplasmic in-
Dobson et al. 2002; A. J. Fry, M. R. Palmer, and D. M. Rand,
unpubl. ms.) and reproduction (e.g., Hoffmann et al. 1990;
Holden, P. R., J. F. Y. Brookfield, and P. Jones. 1993. Cloning and
Snook et al. 2000; Dobson et al. 2002), the interactions we
characterization of an ftsZ cognate from a bacterial symbiont of
found could be important in the interpretation of fitness stud-
Drosophila melanogaster. Mol. Gen. Genet. 240:213–220.
ies using Drosophila and other arthropods that carry Wol-
Hurst, G. D. D., F. M. Jiggins, J. H. G. Von Schulenburg, D. Ber-
bachia. Future work should be directed toward identifying
trand, S. A. West, I. I. Goriacheva, I. A. Zakharov, J. H. Werren,R. Stouthamer, and M. E. N. Majerus. 1999. Male-killing Wol-
chromosomes or chromosomal regions in D. melanogasterbachia in two species of insect. Proc. R. Soc. Lond. B 266:
that are involved in these Wolbachia-induced phenotypes.
Such studies will prove useful for understanding how host
Jeyaprakash, A., and M. A. Hoy. 2000. Long PCR improves Wol-
and parasite genomes coevolve to produce more mutualistic
bachia DNA amplification: wsp sequences found in 76% of sixty-three arthropod species. Insect Mol. Biol. 9:393–405.
Jiggins, F. M., G. D. D. Hurst, and M. E. N. Majerus. 1998. Sex
ratio distortion in Acraea encedon (Lepidoptera: Nymphalidae)
is caused by a male-killing bacterium. Heredity 81:87–91.
Jiggins, F. M., J. K. Bentley, M. E. N. Majerus, and G. D. D. Hurst.
We thank S. Chien, M. Palmer, and L. Sheldahl for assis-
2001a. How many species are infected with Wolbachia? Cryptic
tance in the laboratory. M. Tatar provided access to incubator
sex ratio distorters revealed to be common by intensive sam-
space and assisted with the mortality analyses. We thank S.
pling. Proc. R. Soc. Lond. B 268:1123–1126.
D. Pletcher for providing his WinModEst computer program.
Jiggins, F. M., J. H. G. Von Schulenburg, G. D. D. Hurst, and M.
E. N. Majerus. 2001b. Recombination confounds interpretations
National Science Foundation grant DEB-0104912 to AJF and
of Wolbachia evolution. Proc. R. Soc. Lond. B 268:1423–1427.
grants DEB-9527709 and DEB-9981497 to DMR supported
Miller, P. S., J. Glasner, and P. W. Hedrick. 1993. Inbreeding de-
pression and male mating behavior in Drosophila melanogaster. Genetica 88:29–36.
Min, K. T., and S. Benzer. 1997. Wolbachia, normally a symbiont
of Drosophila, can be virulent, causing degeneration and early
Aspi, J. 2000. Inbreeding and outbreeding depression in male court-
death. Proc. Natl. Acad. Sci. USA 94:10792–10796.
ship song characters in Drosophila montana. Heredity 84:
Olsen, K., T. Reynolds, and A. A. Hoffmann. 2001. A field cage
test of the effects of the endosymbiont Wolbachia on Drosophila
Ballard, J. W. O. 2000. Comparative genomics of mitochondrial
melanogaster. Heredity 86:1–7.
DNA in Drosophila simulans. J. Mol. Evol. 51:64–75.
O’Neill, S. L., R. Giordano, A. M. Colbert, T. L. Karr, and H. M.
Buck, S., M. Nicholson, S. Dudas, R. Wells, A. Force, G. T. Baker
Robertson. 1992. 16S rDNA phylogenetic analysis of the bac-
III and R. Arking. 1993. Larval regulation of adult longevity in
terial endosymbionts associated with cytoplasmic incompatibil-
a genetically-selected long-lived strain of Drosophila. Heredity
ity in insects. Proc. Natl. Acad. Sci. USA 89:2699–2702.
Partridge, L., and K. Fowler. 1990. Non-mating costs of exposure
Caspari, E., and G. S. Watson. 1959. On the evolutionary impor-
to males in female Drosophila melanogaster. J. Insect Physiol.
tance of cytoplasmic sterility in mosquitoes. Evolution 13:
Pletcher, S. D. 1999. Model fitting and hypothesis testing for age-
Chapman, T., L. F. Liddle, J. M. Kalb, M. F. Wolfner, and L.
specific mortality data. J. Evol. Biol. 12:430–439.
Partridge. 1995. Cost of mating in Drosophila melanogaster fe-
Pletcher, S. D., A. A. Khazaeli, and J. W. Curtsinger. 2000. Why
do lifespans differ? Partitioning mean longevity differences in
Stouthamer, R., J. A. Breeuwer, R. F. Luck, and J. H. Werren. 1993.
terms of age-specific mortality parameters. J. Gerontol. 55:
Molecular identification of microorganisms associated with par-
Poinsot, D., and H. Mercot. 1997. Wolbachia infections in Dro-
Turelli, M., and A. A. Hoffmann. 1995. Cytoplasmic incompatibility
sophila simulans: Does the female host bear a physiological cost?
in Drosophila simulans: dynamics and parameter estimates from
natural populations. Genetics 140:1319–1338.
Rousset, F., D. Bouchon, B. Pintureau, P. Juchault, and M. Solignac.
Vaupel, J. W., and A. I. Yashin. 1985. The deviant dynamics of
1992. Wolbachia endosymbionts responsible for various alter-
death in heterogeneous populations. Soc. Methodol. 15:179–211.
ations of sexuality in arthropods. Proc. R. Soc. Lond. B 250:
Werren, J. H., and J. D. Bartos. 2001. Recombination in Wolbachia.
SAS Institute. 1995. JMP statistics and graphics guide. Ver. 3.2
Werren, J. H., and D. M. Windsor. 2000. Wolbachia infection fre-
quencies in insects: evidence for a global equilibrium? Proc. R.
Slatko, B. E., S. L. O’Neill, A. L. Scott, J. H. Werren, and M. L.
Blaxter. 1999. The Wolbachia genome consortium. Microb.
Werren, J. H., D. M. Windsor, and L. R. Gou. 1995a. Distribution
of Wolbachia among Neotropical arthropods. Proc. R. Soc. Lond.
Snook, R. R., S. Y. Cleland, M. F. Wolfner, and T. L. Karr. 2000.
Offsetting effects of Wolbachia infection and heat shock on
Werren, J. H., W. Zhang, and L. R. Gou. 1995b. Evolution and
sperm production in Drosophila simulans: analyses of fecundity,
phylogeny of Wolbachia: reproductive parasites of arthropods. Proc. R. Soc. Lond. B 261:55–63.
fertility, and accessory gland proteins. Genetics 155:167–178.
Yen, J. H., and A. R. Barr. 1971. New hypothesis for the cause of
Solignac, M., D. Vautrin, and F. Rousset. 1994. Widespread oc-
cytoplasmic incompatibility in Culex pipiens. Nature 232:
currence of the proteobacteria Wolbachia and partial cytoplasmic
incompatibility in Drosophila melanogaster. C. R. Acad. Sci. Paris 317:461–470.
These instructions are designed to inform you about Total Knee Replacement. Initial instructions are to be fol owed before the surgery, during your stay in the hospital and after surgery when you are at home. The success of the Total Knee Replacement surgery requires active participation of the patient. For a long-term successful result, understanding and adherence to these guidelines are imper
i>clicker Pedagogy Case Study Lori Morin, PhD College of Health Professions and Biomedical Sciences, School of Pharmacy The University of Montana COURSE OVERVIEW Course Titles/Subject: Pharmacy Practice I and II (Pharmacy 309 and 310) Course Enrollment/Student Information: The courses are taught in the fall and spring semesters, respectively, and each has an enrollment
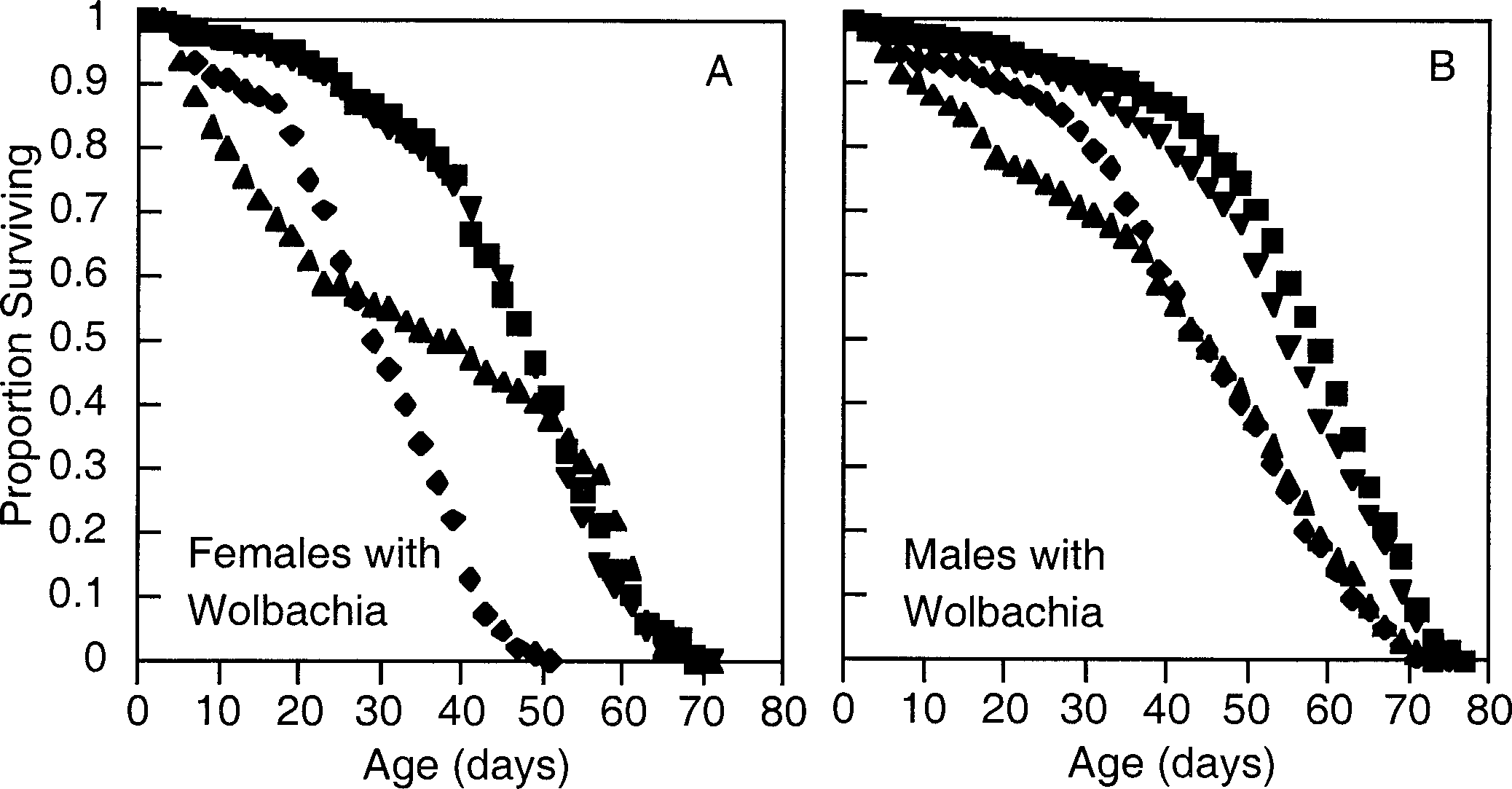
 Survival curves from females (A) and males (B) from
parental crosses and hybrids. Data are from Wolbachia-infected (W)flies reared in mixed-sex mating cages. Symbols: Z53 ϫ Z53, filledtriangle; Z53 ϫ Z2, square; Z2 ϫ Z53, inverted triangle; Z2 ϫ Z2,diamond. (A) Females are significantly heterogeneous by log-rank(LR) test (2 ϭ 254, df ϭ 3, P Ͻ 10Ϫ3), (B) as are males (LR 2
ϭ 270, df ϭ 3, P Ͻ 10Ϫ3).
Survival curves from females (A) and males (B) from
parental crosses and hybrids. Data are from Wolbachia-infected (W)flies reared in mixed-sex mating cages. Symbols: Z53 ϫ Z53, filledtriangle; Z53 ϫ Z2, square; Z2 ϫ Z53, inverted triangle; Z2 ϫ Z2,diamond. (A) Females are significantly heterogeneous by log-rank(LR) test (2 ϭ 254, df ϭ 3, P Ͻ 10Ϫ3), (B) as are males (LR 2
ϭ 270, df ϭ 3, P Ͻ 10Ϫ3).