He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Courses.bio.unc.edu
Chloroquine Transport via the Malaria Parasite’s Chloroquine Resistance Transporter The following resources related to this article are available online at www.sciencemag.org (this information is current as of December 25, 2009 ):
including high-resolution figures, can be found in the online
Updated information and services, version of this article at: Supporting Online Material cites 21 articles subject collections reprints
of this article or about obtaining permission to reproduce this article in whole or in part can be found at:
(print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by the
ScienceAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. Copyright2009 by the American Association for the Advancement of Science; all rights reserved. The title
uptake of [3H]CQ was measured in an acidicmedium (pH = 6.0), in which the majority of CQwas protonated. Oocytes expressing PfCRTCQR
showed a marked (typically 5-fold, and up to 10-fold) increase in [3H]CQ uptake relative to non-
injected controls and to oocytes expressingPfCRTCQS (Fig. 2A). This is consistent withPfCRTCQR, but not PfCRTCQS, mediating the
Rowena E. Martin,1* Rosa V. Marchetti,1 Anna I. Cowan,2 Susan M. Howitt,1
transport of [3H]CQ. (The membrane potential
and cytosolic pH in PfCRTCQS-expressing oocyteswere the same as those in PfCRTCQR-expressing
The emergence and spread of chloroquine-resistant Plasmodium falciparum malaria parasites
oocytes; table S2.) In contrast, oocytes injected
has been a disaster for world health. Resistance is conferred by mutations in the Chloroquine
Resistance Transporter (PfCRT), an integral membrane protein localized to the parasite’s
native (that is, motif-replete, nonharmonized)
internal digestive vacuole. These mutations result in a marked reduction in the accumulation of
PfCRTCQR did not show increased [3H]CQ up-
chloroquine (CQ) by the parasite. However, the mechanism by which this occurs is unclear. We
take, nor was the protein present at significant
expressed both wild-type and resistant forms of PfCRT at the surface of Xenopus laevis oocytes. The
levels in the plasma membrane (fig. S3, A and B).
resistant form of PfCRT transported CQ, whereas the wild-type protein did not. CQ transport via
The sequence modifications made here there-
the mutant PfCRT was inhibited by CQ analogs and by the resistance-reverser verapamil. Thus,
fore facilitated the functional expression of
CQ resistance is due to direct transport of the drug via mutant PfCRT.
T76K and S163R (15) mutations in PfCRTCQR
Malaria,aninfectiousdiseasethatisprev- Thedigestivevacuoleisalysosomalorganelle, restoreCQsensitivitytoCQRparasites(9,16).
and the targeting of PfCRT to this compartment
The introduction of either one of these changes to
is likely to be mediated by discrete endosomal-
PfCRTCQR, each of which entailed the addition
a drug that was cheap, safe, and effective. CQ
lysosomal trafficking motifs. Upon expression of
of a positive charge to the putative substrate-
PfCRT in Xenopus oocytes, such motifs may cause
binding site of the protein (10, 11), resulted in the
falciparum Chloroquine Resistance Transporter
the protein to be targeted to analogous organelles,
loss of CQ transport activity (Fig. 2B). In con-
(PfCRT) (1, 2) and is associated with a marked
rendering direct measurements of PfCRT function
trast, the introduction of K76T to PfCRTCQS did
reduction in CQ accumulation by the parasite
impractical. We therefore identified and removed
not result in a significant increase in [3H]CQ up-
(3, 4). CQ is a diprotic weak base (pKa of 8.1 and
multiple putative trafficking motifs from both
take (Fig. 2B). (PfCRTCQS K76T did localize to
10.2, where Ka is the acid dissociation constant),
termini of the PfCRT protein sequence (fig. S1).
the oocyte plasma membrane; fig. S4.) The K76T
with the relative proportions of the neutral,
In addition, the PfCRT coding sequence for
mutation is therefore necessary but not sufficient
mono-protonated (CQH+), and di-protonated
for the transport of CQ via PfCRT. This is con-
) species varying with pH (table S1).
harmonized to facilitate correct folding of the
sistent with the other PfCRT mutations acting in
The neutral species enters the parasite and its
protein (12, 13). Hemagglutinin (HA)–tagged
synergy with K76T to confer CQ resistance.
internal compartments via simple diffusion.
forms of this modified version of the PfCRT
CQ transport showed a strong dependence on
When the base enters the acidic environment of
sequence were expressed in Xenopus oocytes, and
the pH of the medium (Fig. 2C). Under alkaline
the parasite’s digestive vacuole [pH ~ 5 (5–8)],
conditions, [3H]CQ was taken up to similarly high
the equilibrium is shifted toward the CQH 2+
PfCRT (PfCRTCQS and PfCRTCQR, respectively)
levels in noninjected oocytes and oocytes ex-
species, which is unable to diffuse across the
to the oocyte plasma membrane was confirmed
pressing PfCRTCQR or PfCRTCQS. This is likely
membrane and becomes trapped, thereby accu-
to represent simple diffusion of uncharged CQ
mulating to high concentrations within this com-
The successful expression of (motif-free,
(uptake was nonsaturable at pH = 7.4 and 8.4;
partment. CQ is thought to exert its antimalarial
codon-harmonized) PfCRT at the oocyte surface
fig. S5). In contrast, at pH = 5.0 to 6.9, CQ
effect here by interfering with the detoxification
enabled us to investigate the transport activity of
transport in oocytes expressing PfCRTCQR was
of heme, which is released as a byproduct of
the protein. Except where specified otherwise,
much higher than that in noninjected oocytes or
The key resistance-conferring mutation in
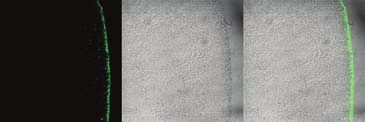
Fig. 1. Immunolocalization of PfCRT in the
Brightfield
PfCRT is the replacement of a lysine (K) with
a threonine (T) at position 76 (9). This K76T
mutation occurs in a region of the protein that
as the pigment layer. This, in turn, sur-
is predicted to be involved in substrate recog-
rounds a cytoplasm crowded with yolk sacs
nition (10). It is never found in isolation, but is
always accompanied by a number of what are
organelles (23). Expression of C-terminally
thought to be compensatory mutations in the pro-
HA-tagged PfCRTCQR or PfCRTCQS results, in
tein (11). We compared the function of mutant
each case, in the appearance of a fluores-
PfCRT from the CQ-resistant (CQR) P. falciparum
indicating that both proteins are expressed
strain Dd2 with that of wild-type PfCRT from
the CQ-sensitive (CQS) strain D10 (fig. S1).
is not present in noninjected oocytes. Simi-
lar results were obtained with N-terminally
Research School of Biology, The Australian National Uni-
versity, Canberra, Australian Capital Territory 0200, Aus-
tralia. 2The John Curtin School of Medical Research, The
Australian National University, Canberra, Australian Capi-
*To whom correspondence should be addressed. E-mail:[email protected]Non-injected
25 SEPTEMBER 2009 VOL 325 SCIENCE www.sciencemag.org
oocytes expressing PfCRTCQS [P < 0.05, analysis
tential undergoing a depolarization over the same
Vmax of 67 T 13 pmol hour−1 per oocyte (Fig. 2E,
of variance (ANOVA)], with the maximum dif-
inset) for the transport of CQ via PfCRTCQR.
ference (that is, the pH optimum for PfCRTCQR-
(The presence of a high extracellular concentra-
mediated CQ transport) observed at pH = 6.0
creased with increasing concentrations of un-
tion of CQ did not affect the membrane poten-
(fig. S6). This pH dependence is consistent with
labeled CQ (Fig. 2E), which is consistent with a
tial or cytosolic pH of noninjected PfCRTCQS- or
CQ being transported in its mono- or di-protonated
saturable transport mechanism. In contrast, raising
forms. Indeed, depolarization of the membrane
the concentration of unlabeled CQ had little ef-
Verapamil increases the accumulation of CQ
potential by the replacement of extracellular Na+
fect on [3H]CQ transport in PfCRTCQS-expressing
by resistant parasites in vitro and thereby increases
with K+ (at pH = 6.0; table S3) resulted in a 25 T
and noninjected oocytes. This is consistent with
their sensitivity to CQ (4). Verapamil inhibited
2% reduction in PfCRTCQR-mediated CQ transport
the entry of the drug into these oocytes being via
the transport of CQ via PfCRTCQR (Fig. 2F and
(Fig. 2D; P < 0.005, paired t test). The decrease
simple diffusion of the neutral species. A least-
Table 1; half-maximum inhibitory concentration
in PfCRTCQR-mediated CQ transport as pH was
squares fit of the data to the Michaelis-Menten
IC50 = 30 T 3 mM), as did a range of quinolines
reduced from 6.9 to 5.5 (Fig. 2C) may have been
equation yielded an apparent Michaelis constant
including quinine and amodiaquine (table S5). In
due, at least in part, to the oocyte membrane po-
KM(CQ) of 245 T 3 mM and a maximum velocity
contrast, piperaquine and artemisinin (both clin-
Time (min) [CQ] (µM) High [K+] [CQ] (µM) [Verapamil] (µM)
Fig. 2. Transport properties of PfCRTCQR in Xenopus oocytes. (A) Oocytes
bars). (D) Effect of depolarization of the oocyte plasma membrane (by
expressing PfCRTCQR (solid circles) showed a marked increase in CQ trans-
replacement of Na+ with K+ in the extracellular medium) on [3H]CQ up-
port relative to noninjected oocytes (solid triangles) and oocytes expressing
take into noninjected oocytes (white bars), oocytes expressing PfCRTCQS
PfCRTCQS (open circles). Rates of CQ uptake (pmol hour−1 per oocyte; n = 3 T
(gray bars), and oocytes expressing PfCRTCQR (black bars). PfCRTCQR-
SEM, estimated from uptake at 60 min) were as follows: noninjected, 1.14 T
expressing oocytes, but not PfCRTCQS-expressing or noninjected oocytes,
0.16; PfCRTCQS-, 1.14 T 0.19; and PfCRTCQR, 5.54 T 0.44. The PfCRTCQR-
showed a significant decrease in CQ uptake when depolarized [P < 0.005
mediated uptake of [3H]CQ (obtained by subtracting uptake in oocytes
and P > 0.05 (ANOVA), respectively]. (E) Effect of unlabeled CQ on the
expressing PfCRTCQR from that in PfCRTCQS-expressing oocytes) was ap-
uptake of [3H]CQ by noninjected oocytes (solid triangles) and oocytes
proximately linear with time for at least 4 hours (inset). (B) Introduction
expressing either PfCRTCQR (solid circles) or PfCRTCQS (open circles). The
of K76T to PfCRTCQS did not increase CQ transport to above that mea-
inset shows the [CQ]-dependence of PfCRTCQR-mediated uptake, which
sured in oocytes expressing PfCRTCQS or in noninjected (ni) oocytes (P >
was calculated by subtracting the uptake measured in oocytes express-
0.05, ANOVA). The introduction of T76K or S163R to PfCRTCQR resulted
ing PfCRTCQS from that in oocytes expressing PfCRTCQR at each CQ
in the loss of PfCRTCQR-associated CQ transport ([3H]CQ uptake in these
concentration. (F) Inhibition by verapamil of the uptake of [3H]CQ by
oocytes did not differ significantly from that in noninjected oocytes or
oocytes expressing PfCRTCQR (solid circles) or PfCRTCQS (open circles). In
from oocytes expressing PfCRTCQS; P > 0.05, ANOVA). (C) pH depen-
all panels, uptake is shown as mean T SEM from three to five separate
dence of [3H]CQ uptake into noninjected oocytes (white bars), oocytes
experiments, within which measurements were made from 10 oocytes per
expressing PfCRTCQS (gray bars) and oocytes expressing PfCRTCQR (black
www.sciencemag.org SCIENCE VOL 325 25 SEPTEMBER 2009
8. T. N. Bennett et al., Mol. Biochem. Parasitol. 133, 99
Table 1. IC50 values for the inhibition of PfCRTCQR-mediated CQ transport by a number of drugs and
peptides. PfCRTCQR-mediated CQ transport was calculated by subtracting the uptake measured in oocytes
9. V. Lakshmanan et al., EMBO J. 24, 2294 (2005).
expressing PfCRTCQS from that in oocytes expressing PfCRTCQR. The data are shown in fig. S7 and Fig. 2F.
10. R. E. Martin, K. Kirk, Mol. Biol. Evol. 21, 1938
IC50 values were derived by least-squares fit of the equation Y = Ymin + [(Ymax – Ymin)/(1 + ([inhibitor]/IC50)C],
where Y is PfCRTCQR-mediated CQ transport, Ymin and Ymax are the minimum and maximum values of Y, and
11. P. G. Bray et al., Mol. Microbiol. 56, 323 (2005). 12. P. Cortazzo et al., Biochem. Biophys. Res. Commun. 293,
C is a constant. All values are mean T SEM from n = 3 or 4 separate experiments, within which mea-
surements were made from 10 oocytes per treatment.
13. A. A. Komar, T. Lesnik, C. Reiss, FEBS Lett. 462, 387
14. Materials and methods are available as supporting
15. Single-letter abbreviations for the amino acid residues
are as follows: F, Phe; G, Gly; H, His; K, Lys; L, Leu; P, Pro;
Q, Gln; R, Arg; S, Ser; T, Thr; V, Val; W, Trp; and Y, Tyr.
16. D. J. Johnson et al., Mol. Cell 15, 867 (2004).
17. D. A. van Schalkwyk, T. J. Egan, Drug Resist. Updat. 9,
18. P. G. Bray et al., Mol. Microbiol. 62, 238 (2006).
19. C. P. Sanchez et al., Biochemistry 44, 9862 (2005). 20. C. P. Sanchez et al., Mol. Microbiol. 64, 407 (2007). 21. B. Naude, J. A. Brzostowski, A. R. Kimmel, T. E. Wellems,
ically effective against both CQS and CQR par-
sensitive) CQ-mediated efflux of H+ from the
22. A. M. Lehane, K. Kirk, Antimicrob. Agents Chemother. 52,
asites) had no effect. Amantadine exhibits some
digestive vacuole of CQR parasites (22). The
antimalarial activity in vitro, particularly against
achievement of a robust expression system for
23. C. A. Wagner, B. Friedrich, I. Setiawan, F. Lang, S. Bröer,
CQR parasites (16), and also inhibited transport
PfCRT has the potential to facilitate the rational
Cell. Physiol. Biochem. 10, 1 (2000).
design of novel CQ-like drugs that bypass the
24. We thank J. Abbey, R. Summers, E. Baker, and R. Slatyer
for technical assistance. This work was supported by the
Several peptides were found to cause a pro-
resistance mechanism and/or the design of clin-
Australian National Health and Medical Research Council
nounced inhibition of CQ transport via PfCRTCQR
ically effective resistance-reversing agents.
(NHMRC) (grant 471472) and the Australian Research
(table S5). Most of the peptides that are active
Council (grant DP0559433). R.E.M. was supported by an
against PfCRTCQR have key elements of the CQ-
NHMRC Australian Biomedical Fellowship (fellowship
resistance reverser pharmacophore [hydrogen bond
1. D. A. Fidock et al., Mol. Cell 6, 861 (2000).
2. A. B. Sidhu, D. Verdier-Pinard, D. A. Fidock, Science 298,
acceptor and two hydrophobic aromatic rings (17)]
(table S6). This pharmacophore can be viewed as
www.sciencemag.org/cgi/content/full/325/5948/1680/DC1
3. C. D. Fitch, Proc. Natl. Acad. Sci. U.S.A. 64, 1181 (1969).
defining the basic elements involved in interactions
4. D. J. Krogstad et al., Science 238, 1283 (1987).
between PfCRTCQR and substrates or inhibitors.
5. R. Hayward, K. J. Saliba, K. Kirk, J. Cell Sci. 119, 1016
The concentration dependence of inhibition
6. N. Klonis et al., Biochem. J. 407, 343 (2007).
of CQ transport was determined for a number of
7. Y. Kuhn, P. Rohrbach, M. Lanzer, Cell. Microbiol. 9, 1004
compounds (Table 1 and fig. S7). YPWF-NH2
(endomorphin-1; an opioid receptor agonist) wasthe most effective peptide inhibitor of PfCRTCQR-
mediated CQ uptake, with an IC50 comparableto that of quinine and verapamil. Measurements
of [3H]YPWF-NH2 uptake in oocytes express-ing different PfCRT constructs revealed that
PfCRTCQR, but not PfCRTCQR-T76K, PfCRTCQR-S163R, or PfCRTCQS, mediates the transport ofthis peptide (fig. S8).
via mutant PfCRT, which provides an explana-
Liam J. Holt,1* Brian B. Tuch,2* Judit Villén,3* Alexander D. Johnson,2
tion for the phenomenon of CQ resistance, as well
as for the reversal of CQ resistance by reversingagents such as verapamil. The presence of a posi-
To explore the mechanisms and evolution of cell-cycle control, we analyzed the position
tive charge (K76 or R163) in the PfCRT substrate-
and conservation of large numbers of phosphorylation sites for the cyclin-dependent kinase
Cdk1 in the budding yeast Saccharomyces cerevisiae. We combined specific chemical inhibition
interacting with the transporter. The K76T muta-
of Cdk1 with quantitative mass spectrometry to identify the positions of 547 phosphorylation
tion removes the positive charge, altering the sub-
sites on 308 Cdk1 substrates in vivo. Comparisons of these substrates with orthologs throughout
strate specificity of PfCRT to allow the transport
the ascomycete lineage revealed that the position of most phosphorylation sites is not
of the protonated drug. In the parasite, the pres-
conserved in evolution; instead, clusters of sites shift position in rapidly evolving disordered
ence of mutant PfCRT on the digestive vacuole
regions. We propose that the regulation of protein function by phosphorylation often
will allow the protonated drug to be transported
depends on simple nonspecific mechanisms that disrupt or enhance protein-protein interactions.
down its electrochemical gradient, out of the vac-
The gain or loss of phosphorylation sites in rapidly evolving regions could facilitate the
uole, and thus away from its site of action (fig. S9).
evolution of kinase-signaling circuits.
This mechanism is consistent with recent studiesimplicating PfCRTCQR in the transport of [3H]CQ
and provide insights into the mechanisms and
in CQR parasites (18–20) and in Dictyostelium
evolution of regulation by phosphorylation. We
discoideum transformants expressing PfCRT at
therefore developed methods for comprehensive
endosomal membranes (21). It is also consistent
tification and analysis of Cdk substrates would
identification of the sites of Cdk1 phosphoryl-
with the recent demonstration of a (verapamil-
enhance our understanding of cell-cycle control
ation on large numbers of substrates in vivo. We
25 SEPTEMBER 2009 VOL 325 SCIENCE www.sciencemag.org
TargeTron Vector pACD4K-C-loxP Catalog Number T2826 Storage Temperature –20 °C TECHNICAL BULLETIN Product Description The TargeTron vector, pACD4K -C-loxP, is a 7,745 bp Escherichia coli expression vector to be used in conjunction with the TargeTron Gene Knockout System, Catalog Number TA0100. This vector differs from the original pACD4K -C vector in that it has lo
¿Qué es el dolor patelofemoral? El dolor patelofemoral es un problema común de la rodilla. Si usted tiene este problema siente dolor debajo y alrededor de su rótula. El dolor puede empeorar cuando usted está activo o cuando se sienta por largo rato. Usted puede tener el dolor sólo en una rodilla o puede tener dolor en ambas rodillas. La causa exacta del dolor patelofemoral se des
 Chloroquine Transport via the Malaria Parasite’s
Chloroquine Transport via the Malaria Parasite’s
 uptake of [3H]CQ was measured in an acidicmedium (pH = 6.0), in which the majority of CQwas protonated. Oocytes expressing PfCRTCQR
showed a marked (typically 5-fold, and up to 10-fold) increase in [3H]CQ uptake relative to non-
injected controls and to oocytes expressingPfCRTCQS (Fig. 2A). This is consistent withPfCRTCQR, but not PfCRTCQS, mediating the
Rowena E. Martin,1* Rosa V. Marchetti,1 Anna I. Cowan,2 Susan M. Howitt,1
transport of [3H]CQ. (The membrane potential
and cytosolic pH in PfCRTCQS-expressing oocyteswere the same as those in PfCRTCQR-expressing
The emergence and spread of chloroquine-resistant Plasmodium falciparum malaria parasites
oocytes; table S2.) In contrast, oocytes injected
has been a disaster for world health. Resistance is conferred by mutations in the Chloroquine
Resistance Transporter (PfCRT), an integral membrane protein localized to the parasite’s
native (that is, motif-replete, nonharmonized)
internal digestive vacuole. These mutations result in a marked reduction in the accumulation of
PfCRTCQR did not show increased [3H]CQ up-
chloroquine (CQ) by the parasite. However, the mechanism by which this occurs is unclear. We
take, nor was the protein present at significant
expressed both wild-type and resistant forms of PfCRT at the surface of Xenopus laevis oocytes. The
levels in the plasma membrane (fig. S3, A and B).
uptake of [3H]CQ was measured in an acidicmedium (pH = 6.0), in which the majority of CQwas protonated. Oocytes expressing PfCRTCQR
showed a marked (typically 5-fold, and up to 10-fold) increase in [3H]CQ uptake relative to non-
injected controls and to oocytes expressingPfCRTCQS (Fig. 2A). This is consistent withPfCRTCQR, but not PfCRTCQS, mediating the
Rowena E. Martin,1* Rosa V. Marchetti,1 Anna I. Cowan,2 Susan M. Howitt,1
transport of [3H]CQ. (The membrane potential
and cytosolic pH in PfCRTCQS-expressing oocyteswere the same as those in PfCRTCQR-expressing
The emergence and spread of chloroquine-resistant Plasmodium falciparum malaria parasites
oocytes; table S2.) In contrast, oocytes injected
has been a disaster for world health. Resistance is conferred by mutations in the Chloroquine
Resistance Transporter (PfCRT), an integral membrane protein localized to the parasite’s
native (that is, motif-replete, nonharmonized)
internal digestive vacuole. These mutations result in a marked reduction in the accumulation of
PfCRTCQR did not show increased [3H]CQ up-
chloroquine (CQ) by the parasite. However, the mechanism by which this occurs is unclear. We
take, nor was the protein present at significant
expressed both wild-type and resistant forms of PfCRT at the surface of Xenopus laevis oocytes. The
levels in the plasma membrane (fig. S3, A and B).