He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Onl_er_ene_2100 1.8
Event-related delta oscillatory responses of Alzheimer patients
G. Yenera, B. Gu¨ntekinb and E. Bas¸arbaDepartments of Neurology and Neurosciences, Brain Dynamics and Multidisciplinary Research Center, Dokuz Eylul University, Izmir,
Turkey; bBrain Dynamics, Cognition and Complex Systems Research Unit, Faculty of Science and Letters, Istanbul Ku¨ltu¨r University,
Background and purpose: Alzheimer type of dementia (AD) is the most common
neuropsychiatric morbidity in elderly individuals. Event-related oscillations (ERO)
provide an useful tool for detecting subtle abnormalities of cognitive processes with
high temporal resolution. Methods: In the present report, event-related oscillations of
patients with AD were analyzed by using a visual oddball paradigm. A total of 22 mildprobable AD subjects according to NINCDS-ADRDA criteria and 20 age-, gender-,
and education-matched healthy control subjects were compared. AD group consisted
from 11 untreated patients and 11 patients treated with cholinesterase inhibitor. Oscillatory responses were recorded from 13 scalp electrodes. Results: Significantdifferences in delta frequency range were seen between the groups by using repeatedmeasures of ANOVA analysis [F(9.120) = 2.228; P = 0.022]. Post-hoc analyses usingWilcoxon test showed that at mid- and left central regions, (Cz, C3) peak amplitudesof delta responses of healthy subjects were significantly higher than either group. Also cholinesterase inhibitors did not have effect on delta oscillatory responses. Conclusions: Our findings imply that the delta oscillatory responses at central loca-tions are highly instable in mild probable AD patients regardless of treatment whencompared to the healthy aged controls. This study supports the importance of oscil-latory event-related potentials for investigating AD brain dynamics.
Event-related theta oscillatory responses have been
proposed to be related to the memory processes [12,17].
One of the leading neurological conditions most
In subjects with ParkinsonÕs disease or schizophrenia,
responsible for neuropsychiatric morbidity in elderly
theta oscillations seem to be less than controls, indi-
individuals is Alzheimer type of dementia (AD). Event-
cating that these oscillations appear to be involved in
related oscillations (ERO) provide a powerful tech-
mnemonic networks [18,19]. Also theta responsiveness
nique, with high temporal resolution, which can be used
in frontal lobes is interpreted as an indication of the
as a tool for detecting subtle abnormalities of cognitive
function of the hippocampo-fronto-parietal system
processes [1,2]. It has been well known for several
during cognitive processes [20,21]. Our recent report
decades that P300 is attenuated in AD. However, the
evaluating phase locking of the visual event-related
full potential of electrophysiological methods in helping
theta oscillations indicated that untreated AD group
to predict [3–5], to diagnose [6–10], and to monitor
has lower phase locking than controls at left frontal
either treatment or progress [11] in AD patients has not
region and the cholinesterase inhibitor treatment in-
been reflected into routine clinical practice.
creases phase locking in theta frequency ranges similar
Event-related oscillatory activity in various frequency
to controls [22]. The question whether cholinergic
bands may reflect different aspects of information
mechanisms affect or modulate event-related oscilla-
processing [1,2]. Alpha oscillatory responses increase
tions in other frequency ranges still remains to be
with simple memory tasks and decrease with demand-
clarified. Investigating these oscillations may help to
ing memory tasks [12,13]. Beta oscillatory responses are
understand differences in brain dynamics of AD
important in attention related tasks in cats and [14],
recognition of facial expression in humans [15,16].
We hypothesized that the AD group would show
lower oscillatory responses than controls. In thisreport, we aimed to compare the peak amplitudes ofevent-related
Correspondence: Go¨rsev G. Yener, M.D., Department of Neurology
frequency ranges in AD subjects, either the untreated
and Neurosciences, Dokuz Eylul University, Izmir 35340, Turkey
or those on cholinesterase treatment, to those of
(tel.: +90 232 412 4050; fax: + 90 232 277 7721; e-mail: gorsev. [email protected]).
Ó 2008 The Author(s)Journal compilation Ó 2008 EFNS
went a cognitive and a complete neurological, neuro-
imaging (CT or MRI) and laboratory examinationincluding blood glucose, electrolytes, liver and kidney
function tests, full blood count, erythrocyte sedimen-
We conducted a prospective open study. Twenty-two
tation rate, thyroid hormone, vitamin B12, HIV,
consecutive, community-dwelling patients suffering
VDRL. Healthy controls were recruited from various
from dementia according to the DSM IV criteria and
community sources; none of them were consanguineous
also with the diagnosis of probable Alzheimer disease
to the patients. The study was approved by the local
according to the NINCDS-ADRDA criteria [23] were
ethics committee. All subjects and relatives gave written
included in the study. AD group was divided into two
groups as the treated and the untreated. In the treatedAD group, eleven subjects (four males, seven females)
were taking only cholinesterase inhibitors (AChEI) as apsychotropic agent for 3–6 months including the titra-
A classical visual oddball paradigm was used in the
tion period (eight subjects were on donepezil 10 mg/day
experiments. Two types of stimuli were used: the stan-
with the initial dose of 5 mg/day that was titrated to
dards and the deviants. The probability of the deviant
10 mg/day by 4 weeks, and three subjects were on riv-
stimuli was 0.20 and that of standard stimuli 0.80. As
astigmine 6–9 mg/day with the initial dose of 3 mg/day,
stimulation we used a white screen with a luminance of
titrated by every 4 weeks either to 6 mg/day or to 9 mg/
35 cd/cm2 for standard signals. The luminance of the
day depending on the tolerance of the drug) and eleven
deviant stimuli were 20% lower (i.e. 28 cd/cm2). The
AD patients (four males, seven females) not taking any
rise-time of the stimulation signal was 10 ms, the
psychotropic medication comprised the untreated AD
duration of the stimulation was 1 second. In all the
group. Both AD groups did not differ from each other
paradigms, the deviant stimuli were embedded ran-
regarding FolsteinÕs Mini-Mental State Examination
domly within a series of standard stimuli. The appli-
(MMSE) scores, ReisbergÕs Global Deterioration Scale
cation of the signal including the rise-fall time and the
(GDS), gender, education, age, or handedness as shown
duration occurs electronically and is supported by a
in Table 1. Time from the onset of symptoms was be-
MATLAB program. Further, the rise time and the dura-
tween one and two years in both AD groups. The
tion of the signal were also checked by means of a
MMSE scores of all AD subjects ranged between 20
photo-sensor recorded in a storage oscilloscope. The
and 24, whereas those of healthy subjects were between
task required was mental counting of the target stimuli.
28 and 30 points. All of the AD subjects were on stage 4
These stimulation signals were applied randomly with
according to the GDS. In treated AD group, the
the inter-stimulus intervals varied between 3 and 7 s.
majority (8 out of 11 subjects) was ÔresponderÕ defined
During the elicitation period of event-related oscilla-
as Ôat least 1 MMSE point increaseÕ 3 months after
tions, all subjects had displayed enough accuracy of
onset of treatment, while three showed decrease in their
mental count of target stimuli, with being slightly worse
scores. Twenty-two healthy elderly control subjects
in both groups of AD than that of controls.
volunteered for the study, two subjects were excludedfor motor artifacts, remaining 20 control subjects
(12 males, 8 females) were not significantly differentfrom both AD groups regarding age, gender, handed-
The EEG was recorded from F3, F4, Cz, C3, C4, T3, T4,
ness and education (Table 1). All AD subjects under-
T5, T6, P3, P4, O1 and O2 locations according to the
SD, standard deviation; NS, non-significant; M, male; F, female; L, left; R, right; GDS,ReisbergÕs Global Deterioration Scale. aChi-square test; bKruskal–Wallis and post-hoc LSDtests.
Journal compilation Ó 2008 EFNS European Journal of Neurology
Event-related oscillation of Alzheimer patients
International 10–20 system. For the recordings an
As oscillatory responses, we measured the peak-to-
EEG-CAP was used. For the reference, EMG and EOG
peak amplitudes of each subjectÕs averaged responses
recordings Ag/AgCl electrodes were used. Linked ear-
filtered in the frequency ranges of delta, theta, alpha,
lobe electrodes (A1 + A2) served as reference. EOG
and beta. The post-stimulus time intervals for peak
from medial upper and lateral orbital rim of the right
amplitudes of oscillatory responses were chosen as
eye was also registered. The EEG was amplified by
follows: frequency ranges of delta and theta, 0–
means of a Nihon Kohden EEG-4421 G apparatus with
600 msn; of alpha and beta 0–250 msn. According to
band limits 0.1–100 Hz 24 dB/octave. The EEG was
the literature of brain oscillations and basic principles
digitized on-line with a sampling rate of 512 Hz and a
of systems theory, the range of oscillatory signals are
total recording time of 2000 ms, 1000 ms of which
chosen in correlation to the frequency signal studied
served as the pre-stimulus baseline.
Computation of selectively averaged ERPs and digital
Statistical Package for the Social Sciences (SPSS) was
Before the averaging procedure, the epochs with arti-
used for statistical analysis. Peak-to-peak maximum
facts were rejected by an off-line technique. In the off-
amplitude responses were separately analyzed for each
line procedure, single sweep EOG recordings were
frequency band by means of a repeated measure
visually studied and trials with eye-movement or blink
ANOVA including the between subjects factor as
artifacts were rejected. Subject averages and grand
groups (healthy aged controls, untreated AD, treated
averages were calculated for each electrode site and
AD) and the within subject factor location (F3, F4, Cz,
experimental condition. The data was digitally filtered
C3, C4, T3, T4, T5, T6, P3, P4, O1 and O2). Greenhouse-
according to determined frequency bands of interest.
Geisser corrected P-values have been taken into con-
In the present study, two approaches were taken in
sideration. Post-hoc analysis was conducted using Wil-
determining the frequency responses of the system: the
transient response frequency characteristics (TRFC)method and digital filtering (DF) method.
Filtering produces visual displays of the time courses
of oscillatory components within the frequency limits of
The peak amplitudes of oscillatory responses in delta,
the utilized filters. The digital filters are advantageous
theta, alpha and beta frequency ranges were measured.
because they do not produce the phase shifts that are a
The only difference in peak amplitudes between groups
characteristic of electronic filters. The digital filtering
was seen in delta oscillations (Table 2).
was employed in the present study for the digital pass-
For the delta frequency range digital filtering was
band filtering of the event-related potentials (ERPs)
determined between 0.5 and 3.5 Hz according to the
and thus to demonstrate the event-related oscillations
AFC. Oscillatory delta responses showed significant
(EROs) in selected frequency-bands (delta: 0.5–3.5 Hz,
differences between event-related responses of healthy
theta: 4–7 Hz, alpha: 8–13 Hz, and beta: 15–30 Hz) [2].
aged controls, untreated and treated AD subjects. The
The numerical evaluation of the frequency charac-
main differences are observed at the central electrode
teristics was accomplished using a Fast Fourier trans-
locations: From the values of Table 3, one can recog-
form (FFT) of the following form: Let Xn be a discrete
nize that in central locations the peak-to-peak ampli-
time series (Xn = X (nDt), T = ((N - 1) Dt). Then the
tudes can be 50–100% larger for controls than for AD
The ANOVA on delta oscillatory responses revealed
[F(9.120) = 2.228; P = 0.022] indicating higher deltaresponse in controls. Post-hoc comparisons using the
where Yk = ak + ibk are the complex Fourier
Wilcoxon paired sample test revealed that the peak-to-
coefficients whose geometric mean is the amplitude
peak delta response was significantly larger for controls
spectrum. According to the results of the amplitude
than for either treated or untreated AD subjects over
frequency characteristics (AFC) the frequencies of
left and mid-central electrodes (P < 0.05 for all com-
interest were determined and the frequency ranges for
parisons) (Figs 1 and 2 and Table 2).
the digital filtering defined. For the frequency ranges
The single sweeps of a typical healthy aged control
grand averages were computed based on single subjectsÕ
subject show good congruence and accordingly phase
averages of the AFCs for each condition and location.
locking of delta responses at Cz, whereas responses of
Ó 2008 The Author(s)Journal compilation Ó 2008 EFNS European Journal of Neurology
Table 2 The mean (SD) peak amplitudes (lV) of oscillatory activities in specific frequency bands in treated AD (t-AD), untreated AD (u-AD) andhealthy elderly controls (Cont). The statistically significant (P < 0.05) results were indicated in bold style
5.10 (1.91) 4.32 (2.32) 4.45 (2.37) 5.27 (2.76) 4.57 (1.90) 5.34 (3.15) 5.05 (2.79) 4.16 (1.53) 3.93 (1.74) 6.19 (2.86) 5.90 (2.09) 8.08 (3.21)
5.02 (2.48) 4.40 (1.60) 4.83 (2.26) 5.76 (2.66) 5.21 (2.55) 5.45 (3.02) 5.74 (3.18) 4.05 (1.83) 3.74 (1.75) 6.56 (2.86) 5.43 (2.09) 7.51 (3.11)
4.01 (1.14) 3.56 (1.27) 3.78 (1.70) 5.53 (3.46) 4.60 (2.06) 5.79 (2.85) 5.74 (3.13) 4.43 (1.05) 4.45 (2.22) 5.52 (1.64) 4.77 (1.93) 8.38 (3.38)
4.28 (2.02) 3.97 (1.27) 3.79 (2.05) 5.64 (3.22) 4.48 (2.04) 5.52 (2.63) 4.97 (2.93) 3.84 (1.09) 3.98 (2.03) 4.48 (2.03) 3.45 (1.76) 7.25 (3.15)
4.40 (1.60) 5.02 (2.48) 4.83 (2.26) 6.12 (2.58) 6.11 (3.09) 5.94 (3.53) 4.74 (2.61) 4.10 (1.73) 4.17 (2.37) 5.97 (2.42) 4.91 (1.06) 7.28 (3.92)
3.85 (1.25) 4.37 (2.55) 3.85 (2.30) 3.42 (2.27) 3.49 (2.06) 3.75 (2.38) 3.00 (1.82) 2.18 (0.69) 2.66 (1.75) 2.94 (1.03) 2.73 (1.15) 4.52 (2.21)
4.90 (2.12) 4.96 (4.01) 3.67 (1.76) 3.97 (1.23) 4.35 (3.38) 4.38 (2.27) 3.15 (1.70) 2.26 (1.40) 2.59 (1.17) 3.96 (2.13) 2.70 (0.85) 4.35 (2.34)
4.05 (1.90) 4.35 (2.05) 4.35 (2.93) 4.14 (1.74) 4.21 (2.09) 3.68 (2.04) 4.39 (2.46) 2.93 (1.10) 3.01 (1.83) 4.40 (1.92) 3.49 (2.11) 5.39 (2.08)
4.34 (2.06) 4.63 (5.77) 4.95 (3.10) 6.87 (6.05) 4.21 (4.17) 5.16 (3.67) 4.40 (2.58) 2.97 (0.55) 3.80 (2.46) 4.19 (1.50) 3.91 (1.57) 5.56 (3.26)
3.67 (0.87) 4.52 (2.29) 4.54 (2.44) 6.39 (3.22) 4.61 (2.60) 5.00 (2.75) 5.94 (1.76) 2.71 (1.76) 4.05 (2.61)
3.63 (1.42) 3.61 (1.36) 4.98 (2.75) 7.02 (4.70) 4.51 (2.62) 5.05 (2.53) 5.04 (1.72) 2.85 (1.20) 4.34 (3.14) 5.00 (1.63) 4.90 (2.33 6.29 (4.37)
O1 4.71 (3.38) 5.45 (2.59) 4.79 (2.82) 5.63 (2.53) 4.65 (2.22) 5.17 (2.84) 6.70 (2.93) 4.39 (2.34) 5.12 (3.28) 8.40 (5.91) 5.74 (2.67) 5.69 (3.25)O2 4.54 (2.04) 4.15 (1.83) 4.87 (2.77) 5.43 (2.03) 4.29 (1.85) 4.98 (2.45) 5.91 (3.60) 4.06 (1.52) 5.20 (3.12) 8.91 (6.34) 5.58 (2.40) 5.92 (3.81)
Table 3 Mean values and standard deviations of delta oscillatory
may play an integrative role in cognitive information
responses and comparisons of the treated AD (t-AD), untreated AD
and in generation of P300 [30]. The possibility of vol-
(u-AD) and healthy elderly controls (Cont) groups with post-hoc
ume conduction from C3 layer of the hippocampus to
the cortex in the generation of P300 is excluded as
animal experiments showed [31]. In human intracranial
recordings, visual working memory task activates
briefly visual association cortex and then activation
soon spreads at once to multiple occipital, parietal andfrontal sites, which all remains active for the entire
C3 7.25 (3.15) 4.48 (2,03) 3.45 (1.76) )2.134 0.033 )2.490 0.013
epoch. Phase-locked oscillations in theta and alpha
8.38 (3.38) 5.51 (1.64) 4.77 (1.93) )2.045 0.041 )2.667 0.008
frequency ranges are prominent in multiple structuresincluding the prefrontal cortex [32].
an AD subject do not (Fig. 1). Phase locking indicates a
Further, according to several authors event-related
stronger response to a given stimulus [25].
potentials arise by superposition of event-related oscil-
In the grand-averages of delta oscillatory responses
lations in various frequency ranges [1]. These ap-
at Cz, it is shown that the control group has larger
proaches hypothesize that the EEG consists of the
amplitude in comparison to either treated or untreated
activity of an ensemble of generators producing oscil-
AD groups (Fig. 2). Peak-to-peak amplitudes of the
latory activity in several frequency ranges. These
control group are 7.25 (3.15) and 8.38 (3.38) lV in C3
oscillators are active usually in a random way. How-
and Cz locations, showing a regular oscillatory pattern.
ever, by application of sensory stimulation these gen-
On the contrary, treated and untreated AD subjects
erators couple and act together in a coherent way.
have smaller amplitudes with an irregular shape
Evoked potentials representing ensembles of neural
population responses were considered as a result oftransition from a disordered to an ordered state [25]. Among event-related oscillations, theta (4–8 Hz) oscil-
lations are correlated with memory load, task difficultyor recognition of previous stimuli [12,13,19,33]. Oscil-
lations at delta frequency range are related to Ôfocused
According to a group of authors, ERPs or P300 re-
attentionÕ, Ôsignal detectionÕ, ÔrecognitionÕ and Ôdecision
sponses are generated in the neocortex, especially in
makingÕ [29,34,35]. In these reports late theta responses
frontal locations [26] or centroparietal/temporoparietal
behave similarly to delta oscillatory responses. Brain
association cortices [27]. Involvement of limbic system
oscillations in lower frequencies are proposed to play a
or hippocampal formation in the generation of P300
role in mediating long range interactions [36]. In
has been also proposed [28,29]. Intracranial recordings
agreement with this, simulation studies have indicated
also suggested that basal ganglia, especially putamen,
that lower frequencies such as delta or theta oscillations
Journal compilation Ó 2008 EFNS European Journal of Neurology
Event-related oscillation of Alzheimer patients
Figure 2 Grandaverages of delta oscillatory response of eachgroup to the target stimuli elicited by a classical visual oddballparadigm recorded from electrodes of C3 and Cz. (a) The healthyelderly control group (n = 20). (b) The untreated Alzheimergroup (n = 11). (c) The treated (cholinesterase inhibitor) Alzhei-mer group (n =11).
oddball paradigms and may be related to signal detec-tion and decision making [20]. As the major shapedetermining oscillatory activity of P300, delta responsesare related to basic information processing mechanismsof attention allocation and immediate memory [39]. Since memory and complex attention functions arehighly reduced in AD [40], our results are in accordancewith cognitive deficits from the psychophysiologicviewpoint.
Topologic distribution and frequency ranges of brain
Earlier reports have shown that the P300 amplitudesare decreased in AlzheimerÕs disease [8,10,39]. Reportson AD using other functional methods such as PET,SPECT or f-MRI also have a tendency to show deficitsat left centro-frontal, left temporo-parietal locations. Arecent report on the event-related oscillatory activity inAD has reported that the significant differences havebeen noted in peak amplitudes of alpha oscillatoryactivity (7–17 Hz) over frontal, central and left tem-poral electrodes [9].
In the present paper, we also found significant dif-
ferences between healthy controls and two groups ofAD subjects in delta oscillatory responses regardless ofcholinergic medication. This difference was insisting
Figure 1 Examples from each group showing single sweeps in
prominently over C3 and Cz in both AD groups in
delta oscillatory frequency range, to the target stimuli elicited by a
classical visual oddball paradigm recorded from the scalp elec-
In our study, classical oddball paradigm was used. In
trode of Cz. The black and thick line indicates the average of single
this task, mental counting of visual target stimuli is
sweeps, and the grey and thin lines show each single sweep for the
considered to be related to memory and complex
subject. (a) An elderly healthy control. (b) An untreated Alzheimer
attention functions. Major reduction in working mem-
subject. (c) A treated (cholinesterase inhibitor) Alzheimer subject.
ory and complex attention observed in Alzheimer pa-tients may be possibly correlated with reduction of
are better suited to sustain long range synchronization
electrical response in central regions. However we
[37]. Not only thalamic neurons but also cortical neu-
cannot exclude the possibility of higher error rates in
rons may discharge in the slow frequency range as delta
detecting target stimuli of AD group may have lead to
[38]. The amplitude of delta response increases during
reduced responses in delta activity.
Ó 2008 The Author(s)Journal compilation Ó 2008 EFNS European Journal of Neurology
Delta difference in left and mid-central positions are
mechanism is yet obscure. Although a great number of
also in accordance with earlier reports of AD subjects
studies establish that both components are physiologi-
studied with fMRI or PET reflecting mainly frontal or
cally separable, we have another argument: This is the
cingulate regions of left hemisphere. Our recent and
selectivity based on the application of a pharmacolog-
other earlier reports imply that in AD, either effects of
ical agent enhancing cholinergic transmission. We also
disease or response to treatment can be more readily
mention here the selectivity of another pharmacological
seen over the left frontal hemisphere.
agent such as valproate with GABAergic or glutama-tergic activity which reduces delta responses in bipolaraffective disorder [44].
Differentiation of delta and theta oscillations in AD
Certainly, it can be stated that in cognitive functions
of Alzheimer patients there is a high decline in working
Earlier functional imaging studies in AD showed that
memory and complex attention [37,45,46]. Since cho-
after administration of AChEI, clinical responders to
linergic medication improves theta phase locking, but
treatment selectively display improvements mainly over
not delta oscillatory response; other transmitters such
left prefrontal areas or left anterior cingulate [41].
as serotonin which increases delta activity in animal
Cortical acetylcholine (ACh) is hypothesized to mediate
studies [47] may help to enhance delta oscillatory
the subjectsÕ abilities to select stimuli and associations
for further processing. The ability of prefrontal cortex
The present paper opens new conjecture to search a
to regulate transmission in more posterior cortical re-
new type of physiological intervention to restore
gions may represent a Ôtop-downÕ mechanism to control
reduction of delta response in central regions. This
attention [42]. Basal forebrain is the main source of
question cannot be answered by this study, but re-
ACh in the neocortex and Alzheimer patients show
depletion in cortical ACh due to degeneration of basal
There are a few conclusions and remarks related to our
forebrain early in the course of illness [43].
findings on AD patients at the initial phase of disease:
We believe that delta oscillatory responses are not
1. Amplitudes of delta oscillatory responses are lower
affected by cholinergic agents, because in our AD
in Alzheimer disease regardless of medication over
group, the degree of clinical impairment did not differ
left and mid-central regions. Activation upon
between the treated and untreated; and subjects in the
treated group was not more advanced than the un-
oscillatory response but not in delta frequency
treated. Finally, majority of the treated group was
considered as responder to treatment. However, a
2. In a way, these two slow oscillatory activities behave
randomized controlled study can give a more probabi-
separately upon application of cholinergic agents.
Possibly, the separation of delta and theta oscilla-
Our recent report evaluating phase locking of the
tory response in AD patients on cholinergic medi-
visual event-related theta oscillations indicated that
cation will gain high importance in future similar
untreated AD group has lower phase locking than
controls at left frontal region. However, the treated AD
3. The studies on event-related oscillations may help
group showed phase locking in theta frequency range
for the diagnostic purposes and also for monitor-
similar to controls [22]. In the present report, peak
ing the effects of pharmacological agents, therefore
amplitudes of delta oscillatory responses are highly re-
in evaluating the transmitter effects.
duced in AD regardless of cholinergic treatment. Therefore cholinergic agents seem to have differentiated
effect on delta and theta responses in AD subjects. Inother words, phase locking in theta oscillatory response
1. Bas¸ar E. EEG-Brain Dynamics. Relation between EEG and
Brain Evoked Potentials. Amsterdam: Elsevier, 1980.
may be sensitive to cholinergic interventions in AD,
2. Bas¸ar E. Memory and Brain Dynamics. Oscillations
whereas amplitudes of delta oscillatory responses are
Integrating Attention, Perception, Learning, and Memory.
One might question whether the separation of delta
3. Babiloni C, Ferri R, Binetti G, et al. Fronto-parietal
and theta responses is a natural way of decomposition.
coupling of brain rhythms in mild cognitive impairment: amulticentric EEG study. Brain Research Bulletin 2006; 69:
If delta and theta responses would behave in a similar
way such a separation could not be strongly assumed.
4. Cichocki A, Shishkin SL, Musha T, et al. EEG filtering
These selective responses to pharmacological agents
based on blind source separation (BSS) for early detection
demonstrate the independent functional correlates of
of AlzheimerÕs disease. Clinical Neurophysiology 2005;
delta and theta responses. However, the underlying
Journal compilation Ó 2008 EFNS European Journal of Neurology
Event-related oscillation of Alzheimer patients
5. Rossini PM, Del Percio C, Pasqualetti P, et al. Conver-
Alzheimer patients treated with cholinesterase inhibitors.
sion from mild cognitive impairment to AlzheimerÕs
International Journal of Psychophysiology 2007; 64: 46–52
disease is predicted by sources and coherence of brain
electroencephalography rhythms. Neuroscience 2006; 143:
23. McKhann G, Drachman D, Folstein M, et al. Clinical
diagnosis of AlzheimerÕs disease: report of the NINCDS
6. Yener GG, Leuchter AF, Jenden D, et al. Quantitative
ADRDA Work Group under the auspices of Department
EEG in frontotemporal dementia. Clinical Electroen-
of Health and Human Services Task Force on AlzheimerÕs
Disease. Neurology 1984; 34: 939–44.
7. Babiloni C, Benussi L, Binetti G, et al. Genotype (cystatin
24. Bas¸ar E. Brain Function and Oscillations: I. Brain Oscil-
C) and EEG phenotype in Alzheimer disease and mild
lations. Principles and Approaches. Heidelberg, New York:
cognitive impairment. Neuroimage 2006; 29: 9948–964.
8. Polich J, Herbst KL. P300 as a clinical assay: rationale,
25. Yordanova J, Kolev V. Single sweep analysis of the theta
evaluation, and findings. International Journal of Psycho-
frequency band during an auditory oddball task. Psy-
9. Karrasch M, Laine M, Rinne JO, et al. Brain oscillatory
26. McCarthy G, Wood CC. Scalp distributions of event–re-
responses to an auditory-verbal working memory task in
lated potentials: an ambiguity associated with analysis of
mild cognitive impairment and AlzheimerÕs disease.
variance models. Electroencephalography and Clinical
International Journal of Psychophysiology 2006; 59: 168–
Neurophysiology 1985; 62: 203–208.
27. Verleger R, Heide W, Butt C, et al. Reduction of P3b in
10. Jeong J. EEG dynamics in patients with AlzheimerÕs dis-
patients with temporo-parietal lesions. Cognitive Brain
ease. Clinical Neurophysiology 2004; 115: 1490–1505.
11. Jelic V, Johansson SE, Almkvist O, et al. Quantitative
28. Wood CC, Allison T, Goff WR, et al. On the origin
electroencephalography in mild cognitive impairment:
of P300 in man. Progress in Brain Research 1980; 54:
longitudinal changes and possible prediction of Alzhei-
merÕs disease. Neurobiology Aging 2000; 21: 533–540.
29. Halgren E, Smith ME. Cognitive evoked potentials as
12. Jensen O, Tesche CD. Frontal theta activity in humans
modulatory processes in human memory formation and
increases with memory load in a working memory task.
retrieval. Human Neurobiology 1987; 6: 129–139.
European Journal of Neurosciences 2002; 15: 1395–1399.
30. Rektor I, Bares M, Kanovsky P, et al. Cognitive poten-
13. Gevins A, Smith ME, McEvoy L, et al. High resolution
tials in the basal ganglia-frontocortical circuits. An
EEG mapping of cortical activation related to working
intracerebral recording study. Experimantal Brain Re-
memory: effects of task difficulty type of processing, and
practice. Cerebral Cortex 1997; 7: 374–385.
31. Bas¸ar-Eroglu C, Bas¸ar E. A compound P300-40 Hz re-
14. Wrobel A. Beta activity: a carrier for visual attention.
sponse of the cat hippocampus. International Journal of
Acta Neurobiologiae Experimentalis 2000; 60: 247–260.
15. Gu¨ntekin B, Bas¸ar E. Emotional face expressions are
32. Halgren E, Boujon C, Clarke J, et al. Rapid distributed
differentiated with brain oscillations. International Journal
fronto-parieto-occipital processing stages during working
of Psychophysiology 2007; 64: 91–100 (e-pub: 5 December
memory in humans. Cerebral Cortex 2002; 12: 710–728.
33. Klimesch W, Hanslmayr S, Sauseng P, et al. Oscillatory
16. O¨zgo¨ren M, Basar-Eroglu C, Bas¸ar E. Beta oscillations in
EEG correlates of episodic trace decay. Cerebral Cortex
face recognition. International Journal of Psychophysiol-
34. Stampfer HG, Bas¸ar E. Does frequency analysis lead
17. Klimesch W, Doppelmayr M, Pachinger T, et al. Brain
to better understanding of human event related poten-
oscillations and human memory performance: EEG cor-
tials. International Journal of Neuroscience 1985; 26:
relates in the upper alpha and theta bands. Neuroscience
35. Demiralp T, Bas¸ar E. Theta rhythmicities following ex-
18. Schmiedt C, Meistrowitz A, Swendemann G, et al. Theta
pected visual and auditory targets. International Journal of
and alpha oscillations reflect differences in memory
Psychophysiology 1992; 13: 147–160.
strategy and visual discrimination performance in patients
36. von Stein A, Sarnthein J. Different frequencies for dif-
with ParkinsonÕs disease. Neuroscience Letters 2005; 388:
ferent scales of cortical integration: from local gamma to
long range alpha-theta synchronization. International
19. Schmiedt C, Brand A, Hildebrandt H, et al. Event-related
Journal of Psychophysiology 2000; 38: 301–313.
theta oscillations during working memory tasks in pa-
37. Kopell N, Ermentrout GB, Whittington MA, et al.
tients with schizophrenia and healthy controls. Cognitive
Gamma rhythms and beta rhythms have different syn-
chronization properties. Proceedings of the National
20. Bas¸ar-Erog˘lu C, Bas¸ar E, Demiralp T, et al. P300-re-
Academy of Sciences USA 2000; 97: 1867–1872.
sponse: possible psychophysiological correlates in delta
38. Steriade M, Gloor P, Llinas RR, et al. Basic mechanisms
and theta frequency channels. A review. International
of cerebral rhythmic activities. Electroencephalography
Journal of Psychophysiology 1992; 13: 161–179.
and Clinical Neurophysiology 1990; 76: 481–508.
21. Demiralp T, Bas¸ar-Erog˘lu C, Rahn E, et al. Event-related
39. Polich J, Kok A. Cognitive and biological determinants of
theta rhythms in cat hippocampus and prefrontal cortex
P300: an integrative review. Biological Psychology 1995;
during an omitted stimulus paradigm. International
Journal of Psychophysiology 1994; 18: 35–48.
40. Cummings JL, Miller BL, Hill MA, et al. The neuropsy-
22. Yener GG, Gu¨ntekin B, O¨niz A, et al. Increased frontal
chiatric aspects of multi-infarct dementia and Alzheimer
type. Archives of Neurology 1987; 44: 389–393.
Ó 2008 The Author(s)Journal compilation Ó 2008 EFNS European Journal of Neurology
41. Mega MS, Dinov ID, Porter V, et al. Metabolic patterns
stimuli in a group euthymic bipolar patients in comparison
associated with the clinical response to galantamine
to healthy controls. 62nd Annual Scientific Convention and
therapy. Archives of Neurology 2005; 62: 721–728.
Program, May 17–May 19, 2007, San Diego, California,
42. Sarter M, Hasselmo ME, Bruno JP, et al. Unraveling the
attentional functions of cortical cholinergic inputs: inter-
45. Miller BL, Read SL, Mahler ME, et al. Altered mental
actions between signal- driven and cognitive modulation
status in the elderly. Primary Care 1984; 11: 653–665.
of signal detection. Brain Research Reviews 2005; 48: 98–
46. Mesulam M-M. Patterns in behavioral neuroanatomy;
association areas, the limbic system, and hemispheric spe-
43. Perry EK, Irving D, Kerwin JM, et al. Cholinergic
cialization. In: Mesulam M-Med. Principles of Behavioral
transmitter and neurotrophic activities in Lewy body
Neurology. Philedelphia, F. A. Davis, 1985: 1–70.
dementia: similarity to ParkinsonÕs and distinction from
47. Schu¨tt A, Bas¸ar E. The effects of acetylcholine, dopamine
Alzheimer disease. Alzheimer Disease and Associated
and noradrenaline on the visceral ganglion of Helix
pomatia. II. Stimulus evoked field potentials. Comparative
44. O¨zerdem A, Kocaaslan S, Tunca Z, et al. Effect of val-
Biochemistry and Physiology – Part C: Toxicology and
proate on oscillatory delta frequency responses to visual
Journal compilation Ó 2008 EFNS European Journal of Neurology
HERBAL STRATEGIES FOR THE PRE- AND POST- OP BODY Herbs help the body heal faster, minimize scaring, maximize sensation, and lessen nervedamage to surgically altered bodies. Lay out a timeline for when to begin preparing foroptimal healing post-surgically. We will discuss herb and drug interactions, includinghormones and anesthesia. This class is also geared toward western practitioners who w
LAS EMOCIONES Lección 1 Para el 1º de enero de 2011 Sábado 25 de diciembre LEE PARA EL ESTUDIO DE ESTA SEMANA: 2 Samuel 13; Gálatas 5:22; Colosenses 3:12-14; Lucas 19:41-44; Juan 16:20-24. PARA MEMORIZAR: “De cierto, de cierto os digo, que vosotros lloraréis y lamentaréis, y el mundo se alegrará; pero aunque vosotros estéis tristes, vuestra tristeza se convertirá en
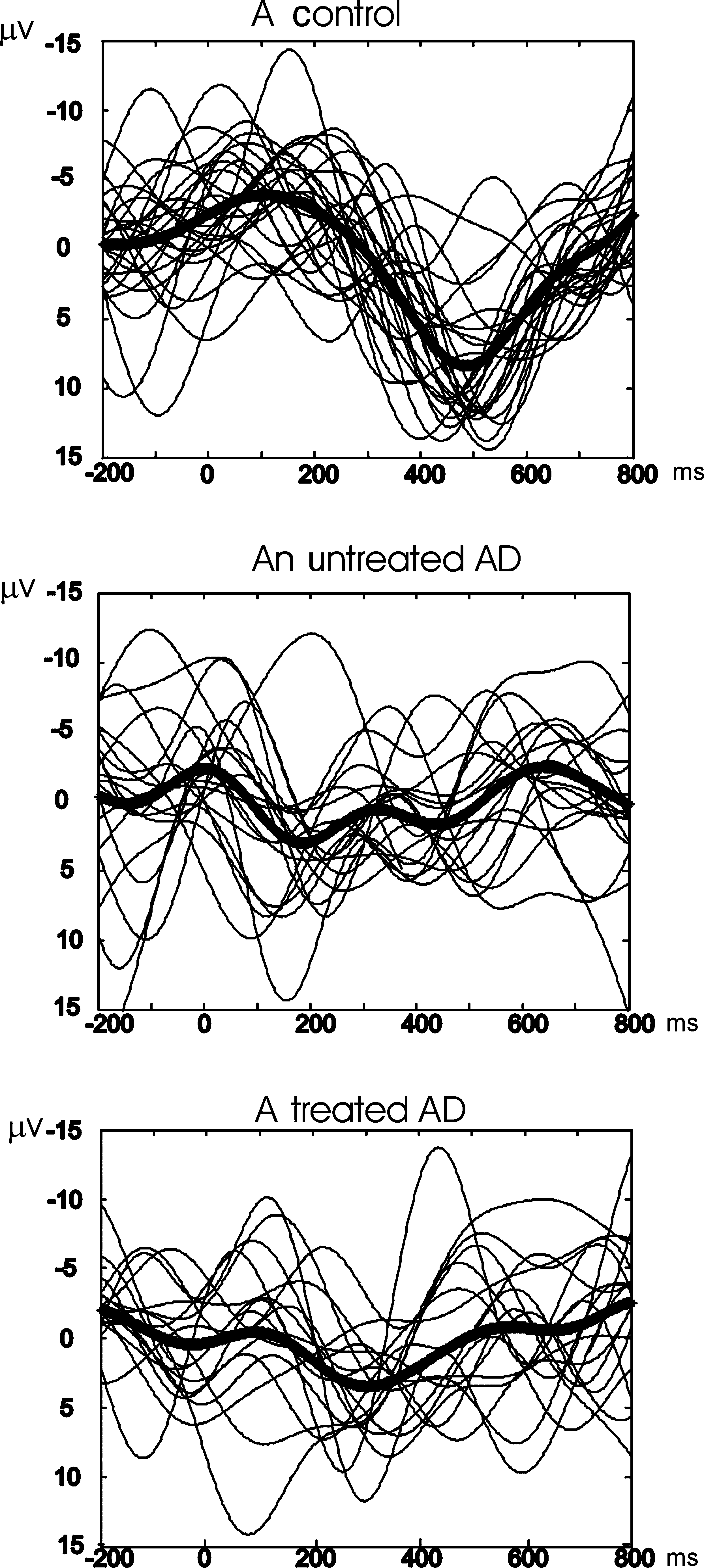
 Event-related oscillation of Alzheimer patients
Figure 2 Grandaverages of delta oscillatory response of eachgroup to the target stimuli elicited by a classical visual oddballparadigm recorded from electrodes of C3 and Cz. (a) The healthyelderly control group (n = 20). (b) The untreated Alzheimergroup (n = 11). (c) The treated (cholinesterase inhibitor) Alzhei-mer group (n =11).
Event-related oscillation of Alzheimer patients
Figure 2 Grandaverages of delta oscillatory response of eachgroup to the target stimuli elicited by a classical visual oddballparadigm recorded from electrodes of C3 and Cz. (a) The healthyelderly control group (n = 20). (b) The untreated Alzheimergroup (n = 11). (c) The treated (cholinesterase inhibitor) Alzhei-mer group (n =11).