He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Talassemiaricerca.unife.it
The use of cell culture procedures for studying fetal hemoglobin stimulating drugs High fetal hemoglobin (HbF) has been shown to ameli- orate the clinical symptoms of patients with β M Hadassah University Hospital, Jerusalem, Israelglobinopathies, β-thalassemia (β-thal) and sickle cell anemia (SCA). Research is therefore focus on finding drugs capable of reactivating the γ-globin genes and stimulating the production of HbF. Several in vitro experimental models have been developed to serve this purpose. Two models are the subject of this review: in
ical symptoms of the underlying disease. Theoretical
vitro established erythroid-like cell lines and primary cul-
considerations 3-5 suggested that in SCA not only do
tures of erythroid cells derived from progenitors
HbF-containing cells have a lower concentration of
obtained from nor mal donors and patients with β-thal
sickle Hb, but HbF inhibits polymerization of this Hb
and SCA. These experimental models are useful for
directly, accounting for the lower propensity of such
cells to undergo sickling. In β-thal elevated HbF shouldcompensate partially for the deficiency in α-chains. These studies ar
These prompted the search for drugs that can ele-
fective and safe drugs. eening of compounds and for studying their mech- anism of action at the cellular and molecular levels.G e essential for finding, testing and devel- oping new, ef
vate γ-globin synthesis and HbF production, which
KEY WORDS:
Erythroid precursors - Fetal hemoglobin - β-tha-
yielded the drug hydroxyurea (HU) that has been
shown to improve the clinical condition in SCA.6, 7Trials with HU have also been carried out in thal
The β-hemoglobinopathies, the Y
β-thalassemias patients with considerable success. Although HU is rel-
atively safe, it is a myelo-suppressive drug. Moreover,
-thal) and sickle cell anemia (SCA) are the most
common class of inherited single-gene diseases.
about 30% of the patients do not respond to HU.
Although important advancement has been made in
Thus, more effective and less hazardous drugs are
prenatal diagnosis and the symptomatic treatment of
the diseases (such as blood transfusions and iron che-
Several HbF-stimulating agents have been discov-
lation), these diseases still have a devastating impact
ered as a result of clinical observations. Thus, buty-
on the affected individuals, their families and the com-
rates were discovered following observations of a delayin HbF disappearance in infants of diabetic mothers 8
munities where the diseases are prevalent. The devel-
and of the increased HbF in patients with urea-cycle dis-
opment of new therapies is, therefore, urgently need-
orders receiving sodium 4-phenylbutyrate.9 However,
ed. Epidemiological studies 1, 2 have shown that high
in order to systematically test HbF-stimulating agents
levels of fetal hemoglobin (HbF) ameliorate the clin-
experimental models are needed. These include invitro established, immortalized, cell lines that can be
Address reprint requests to: Prof. E. Fibach, Department of Hematology,
induced to express some erythroid characteristics and
Hadassay University Hospital, Ein-Kerem, Jerusalem 91120, Israel. E-mail: [email protected]
primary cultures of erythroid progenitors.
CULTURE MODELS FOR STUDYING HBF STIMULATION
Established erythroid-like cell lines
or clusters in suspension.20-22 In both systems EPO isessential for full development of Hb-containing eryth-
Human erythroid-like cell lines, such as K562,10
HEL 11 and UT-7,12 were derived from cells explantedfrom patients with various forms of myeloid leukemia. Semi-solid cultures A
The cells were adapted to grow in culture and even-tually became immortalized. They grow as single,
In the cloning method, suspensions of single hemat-
undifferentiated, cells in suspension, with little produc-
opoietic cells derived from the bone marrow, periph-
tion of Hbs. When stimulated by various agents they
eral blood or other sources such as fetal liver, neona-
respond within a few days with a significant increase
tal cord blood or adult spleen, are dispersed in semi-
in the production of Hbs and the expression of some
solid media usually containing methyl cellulose or
other erythroid specific differentiation markers.13 Thus,
plasma clot. Colonies start to appear after 3-4 day
the K-562 cell line can be induced by agents, such as
incubation, and they reach their final size and hemo-
hemin, 5-azacytidine, HU, butyroids and other agents
globinization after 1-2 weeks. Each colony represents
to synthesize embryonic Hbs (Gower 1, Gower 2 and
a clone derived from one erythroid committed progen-
Portland) and HbF.14-18 These cell lines provide repro-
itor. Based on the final size of the colonies and the
ducible, uniform, large populations of cells that under-
time required for their hemoglobinization various
go a rather synchronized differentiation program.
types of progenitors can be distinguished; the late
Although cell lines serve as convenient experimental
erythroid colony forming units (CFUe) reach final size
models, because of their leukemic origin and long
and hemoglobinization after 1 week and then disap-
history in culture, they do not recapitulate all aspects
pear, while the early erythroid burst forming units
of erythropoiesis. For example, K562 cells do not
A (BFUe) develop after 2 weeks.23, 24Erythroid colonies
response to erythropoietin (EPO), the physiological
can be distinquished from other (myeloid) colonies by
inducer of erythroid cell proliferation and differentia-
their red color or by their positive reaction with heme-
tion. In addition, they do not produce adult Hbs (HbA
specific reagents.25 Counting the different types of
colonies on different days provides a quantitative esti-
2). In essence, stimulation of these cells
involves increase production of Hb types that are pro-
mation of the frequency of the various progenitors
duced constitutively at background levels when un-
in the hematopoietic tissue. Cellular analysis of the cul-tures can be performed on the entire cell populationby harvesting the culture and washing out the meth-
stimulated, rather than their reactivation from a non- active to an active state. Moreover, in the absence ofG
-globin, this model does not recapitulate
ylcellulose or by picking up (with a capillary tube)
individual colonies and then pooling them together.
of α-chains). And finally, although many agents have
Alternatively, the analysis can be performed on single
been shown to increase Hb production in such cell
colonies after they were picked up individually.
Analysis of the Hb content of individual colonies rep-
cultures, and, vice versa, other agents (e.g., cytokines)
resents their heterogeneity with respect to their poten-
fect primary cultures fail to stimulate cell lines.
tial to produce HbF. When single cell techniques areused, such as staining cells with specific antibodies, theheterogeneity among progeny of individual progen-
Primary cultur es of human erythroid cells
itors can be assessed. This model has been used for
The establishment of a cell line is a rare event; nor-
The fact that in this culture system the cells are
mal cells and cells from most patients cannot be
immobilized in semi-solid medium results in several
immortalized. Nevertheless, primary cultures of eryth-
disadvantages. The cell yield per colony and the total
roid cells can be readily established from most normal
cells per culture are low (<105/ml) making it techni-
individuals and patients. Their growth in vitro repre-
cally difficult to carry out biochemical, molecular and
sents more closely than cell lines the in vivo situa-
immunological characterizations of the developing
tion. The cells can be cultured either in semi-solid
cells. It is a one-step continuous culture and addition
medium where they develop into discrete colonies,19
of the tested agents during the culture is difficult. In
or in liquid medium where they grow as single cells
addition, the proportion of HbF produced in colo-
CULTURE MODELS FOR STUDYING HBF STIMULATION
nies grown in semi-solid medium is significantly high-er than that produced in vivo by the donor of the
The liquid culture procedure overcomes several of
these obstacles. It is possible to obtain large cultures
of relatively pure and synchronized erythroid cell
Culture in liquid medium+fetal bovine serum
population and compounds can be added on differ-
+ erythropoietin
ent days when the culture consists of cells at specif-
Liquid medium+fetal bovineserum+cytokine-containing
ic stages of maturation. In a procedure developed by
us the culture is divided into 2 phases (Figure 1): an
EPO-independent phase, in which peripheral blood
cells are first cultured in the presence of a combina-tion of growth factors, but in the absence of EPO,where early erythroid committed progenitors, BFUe,
EProliferation and Proliferation and maturation differentiation into Hb-containing erythroid
proliferate and differentiate into CFUe-like progenitors. of BFU into CFUe
phase, the latter cells, cultured in an EPO-
supplemented medium, continue to proliferate and
Figure 1.—A schematic flow chart of the 2-phase liquid culture pro-
mature into orthochromatic normoblasts and enu-
In phase I of the procedure, peripheral blood
mononuclear cells isolated on Ficoll or an equiva-
cultures contain some adherent cells (mainly macro-
lent 1.077 g/ml density medium are cultured in liquid
phages) and non-adherent cells (mainly lymphocytes).
medium supplemented with human recombinant
The non-adherent cells are harvested, washed and
, interleukin-6 and stem cell factor.
re-cultured in fresh medium supplemented with EPO
These cytokines can be replaced by conditioned
(1 U/ml). In the absence of necessary cytokines to
medium derived from cultures of various human car-
support their proliferation and differentiation, non-
cinoma cell lines, such as the 5 637 bladder carcino-G
erythroid progenitors cease their development.
ma. This conditioned medium contains a variety of
Erythroid progenitors (CFUe), following exposure to
EPO, proliferate and differentiate into erythroid pre-
culture is, therefore, EPO-independent. Lymphocytes
cursors. Proerythroblasts are discernible by inverted
can be removed from the inoculum using specific
microscopy on days 4-5 of phase II as large, round
and smooth cells. At this stage they may be purified
lymphocytic (CAMPATH-1) antibodies 31 or by sup-
from remaining lymphocytes and erythrocytes on
pressing their activation and proliferation by addi-
Percoll gradient (1.0585 g/ml) and re-cultured in the
same medium. As the proerythroblasts continue to
During phase I, erythroid, myeloid and megakaryo-
multiply, they form clusters and then large aggregates
cytic progenitors proliferate and differentiate. The
which when undisturbed can reach hundreds of cells
early erythroid-committed progenitors, BFUe, prolife-
(Figure 2A). As these cells differentiate they decrease
rate and differentiate into CFUe-like progenitors. The
in size and accumulate Hb and the aggregates assume
kinetics of expansion of the erythroid progenitors in
phase I of the culture and their requirement for cyto-
Using this procedure, erythroid cells continue to
kines were studied by direct and indirect cloning in
proliferate in phase II for 14 days; their number peaks
on day 12 and reaches up to 3x106 cells/ml culture;
Following 1-week incubation in phase I further
more than 96% of which are Hb-containing cells -
development of CFUe requires EPO; in its absence,
mostly (90%) orthochromatic normoblasts and 4%
these cells are blocked and synchronized with respect
enucleated erythrocytes (Figures 2B, C). Non-erythroid
to differentiation and proliferation. At this stage, the
cells usually constitute less than 4% of the total pop-
CULTURE MODELS FOR STUDYING HBF STIMULATION
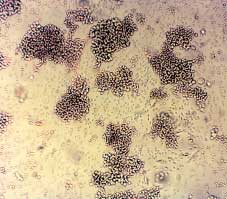
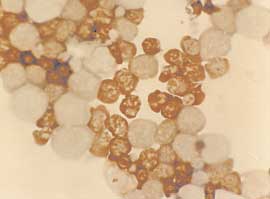
Figure 2.—Erythroid cells in culture. A) Lar
ge aggregates of erythroid precursors on day 6 of phase II. Unstained cells were photographed
in situ using an inverted microscope. B) Benzidine staining of cells from day 8 phase II cultures. Cells were smeared on a glass slide using
a cytocentrifuge, and stained with benzidine. Mature erythroid Hb-containing cells which are benzidine positive (B+) and early erythroid pre-cursors which are benzidine negative (B-) are seen. C) Benzidine/Giemsa staining of cells from day 10 cultures. Cells were smeared as in Bfollowed by staining with Giemsa. Both B+ erythroid cells and B- myeloid and lymphoid cells are seen. D) Cells harvested on day 12 of theculture.
ulation. Following the beginning of maturation, eryth-
blood from patients with thal or SCA, because of the
roid cells can be purified to >99% using immuno-
high frequency of erythroid progenitors,23 20 ml of
magnetic beads directed against erythroid specific
whole blood can be used for 20 ml cultures that even-
membrane antigen such as glycophorin A.32 For nor-
mal erythroid cultures, the buffy coat fraction (that is
Peripheral blood cells are employed in this proce-
usually discarded by the Blood Bank) of a whole
dure for the following reasons: a) the availability of PB
blood unit may be used for setting up 100 ml cul-
from normal individuals and patients; b) the homoge-
tures. In such cases, a total of up to 3×108 cells can be
neity of the peripheral blood erythroid progenitors,
harvested on day 12 of the culture (Figure 2D). With
namely early BFUe, as opposed to the BM which con-
CULTURE MODELS FOR STUDYING HBF STIMULATION
tains progenitors at various developmental stages.33
Good results can be obtained with cells derived fromother sources, including CD34+ cells purified by immu-
no-magnetic bead technologies, but, in these cases
some modifications of the procedure are required.
This system recapitulates many aspects of in vivo
erythropoiesis including globin RNA metabolism,34-36
cell cycle kinetics,37 cell surface antigens,38 iron andferritin metabolism 39-42 and transcription factors.37, 43
We have used this system to study the effects of
hundreds of compounds, including butyroids,44, 45hemin 46 and EPO 47 and histone deacetylase inhibi-
tors.48 For studying their potential to enhance HbFproduction, compounds can be added to phase I,
phase II or both. Non-toxic drugs such as cytokines
and hemin may be added to the cultures at any time. With cytotoxic drugs, such as HU and 5-azacytidine,
because of their cyto-toxic/-static effects, they are
usually added on day 4-8 of phase II.
Since the erythroid cells in phase II are grown in
suspension, samples of cells can be withdrawn at any
time without disturbing the cultures and assayed for
sis, cell cycle or expression of surface antigens.
Hemoglobinization can be easily followed by staining
Figure 3.—HPLC chromatogram of hemoglobins produced by cul-
the cells with benzidine solutions.25 The benzidine
tured erythroid cells. Cells were harvested from day 12 cultures,washed and lysed. The hemoglobins in the lysate were separated on
staining is specific for heme-containing compounds.
cation-exchange HPLC. The peaks are labeled with the retention timesand the corresponding hemoglobin type.
Smear of cells on a glass slide can be stained with a combination of 3’,-3’,-dimethoxybenzidine, whichG
stains the Hb-containing cells brown, and Giemsa,which permits detailed examination of the cell mor-
The distribution of the erythroid cell population
phology (Figures 2B, C). The “blue” method with
with respect to intracellular content of HbF can be
dihycrochloric benzidine stains the cells in their orig-
analyzed by flow cytometry using monoclonal antibod-
inal medium. It requires one drop of culture and after
ies directed specifically against HbF 52, 53 (Figure 4).
1 minute the cells are scored in a hemocytometer and
Dual/triple staining with the corresponding antibod-
the number and percentage of positive cells deter-
ies can be used for simultaneous analysis of both
mined. Using this procedure, positive cells can be
HbA (or HbS) and HbF, or Hb and another marker of
routinely found after 4-5 days in phase II.
interest such as glycophorin A or CD36 surface anti-
The Hb content of the developing erythroid cells
can be measured by a variety of techniques, such as
The results with HU indicated that cultured cells
the alkaline denaturation and benzidine staining 49
recapitulated many aspects of hemopoiesis in HU-
and high-performance liquid chromatography (HPLC),
treated patients: a reduction in cell number, an increase
cation-exchange HPLC for hemoglobins (Figure 3) and
in cell size (mean corpuscolar volume, MCV), an
reverse-phase HPLC for globin chains.50, 51 Using the
increase in total Hb per cell (mean corpuscolar hemo-
HPLC techniques, Hb is measurable in culture as ear-
globin, MCH) and an increase in the proportion of
ly as 5 days in phase II. On day 12, one ml culture is
HbF.54 Using cultures derived from SCA patients, we
sufficient for multiple measurements. The mean cellu-
have shown that following treatment with HU and
lar Hb or HbF concentration of erythroid cells are cal-
other HbF-stimulating drugs, sickle Hb polymerization
culated from the values of the HPLC determinations
and cell sickling in low oxygen atmosphere was inhib-
divided by the number of benzidine-positive cells.
ited.55 Moreover, preliminary data suggest a correlation
CULTURE MODELS FOR STUDYING HBF STIMULATION
M T Glycophorin A
Figure 4.—Flow cytometry of fetal hemoglobin in cultured thalassemic cells. HU (150 µM) untreated and treated cultures derived from a β-thalassemia patient were stained with PE-conjugated antibodies for HbF and with FITC-conjugated antibodies for glycophorin A. In untreat-ed cultures 12.6% of the erythroid cells (glycophorin A positive) were also positive for fetal hemoglobin (cells in the upper right quadrant). In HU-treated cultures 24.0% of the erythroid cells were positive for fetal hemoglobin.
suited for growing erythroid cells from normal and
to treatment. If this correlation between the in vitro
patients. It yields large, pure and synchronized (in
results is substantiated, it will be possible
terms of maturation) erythroid populations. This
by testing cultures of cells derived from the patient's
system recapitulates the in vivo pattern of Hb pro-
peripheral blood to predict the response of the patient
duction of the donors. Following evaluation of drugs
in vivo. This will prevent both expensive and poten-G
in these in vitro systems, the most promising drugs
tially risky treatment from patients who do no respond
should then be further studied in animal model
systems, such as transgenic mice and in primates
Conclusions References
1. Haghshenass M, Ismail-Beigi F, Clegg JB, Weatherall DJ. Mild sick-
le-cell anaemia in Iran associated with high levels of fetal haemog-
tion in β-thal and SCA. The in vitro models aim main-
2. Al-Awamy BH, Niazi GA, el-Mouzan MI, Altorki MT, Naeem MA.
ly for: A) high throughput assay for testing compounds
Relationship of haemoglobin F and alpha thalassaemia to sever-
for HbF-stimulating potential. Erythroid cell lines,
ity of sickle-cell anaemia in the Eastern Province of Saudi Arabia. Ann Trop Paediatr 1986;6:261-5.
such as K562, can serve nicely this purpose. B) Cellular
3. Benesch RE, Edalji R, Benesch R, Kwong S. Solubilization of hemo-
and molecular study of their mechanism of action.
globin S by other hemoglobins. Proc Natl Acad Sci U S A 1980;77:
For this purpose, the model should be as similar as
4. Noguchi CT, Rodgers GP, Serjeant G, Schechter AN. Levels of fetal
possible to the cells of interest, namely, normal, β-thal
hemoglobin necessary for treatment of sickle cell disease. N Engl
and SCA cells. C) Develop a predicting assay for an
5. Eaton WA, Hofrichter J. The biophysics of sickle cell hydroxyurea
individual patient’s response to a particular drug or
drug combination. For this purpose the assay should
6. Charache S, Terrin ML, Moore RD, Dover GJ, Barbon FB, Eckert SV
use cells obtained from the patient under study. et al. Effect of hydroxyurea on the frequency of painful crises in sick-le cell anemia. Investigators of the Multicenter Study of Hydroxyurea
The 2-phase liquid culture procedure is especially
in Sickle Cell Anemia. N Engl J Med 1995;332:1317-22.
CULTURE MODELS FOR STUDYING HBF STIMULATION
7. Steinberg MH, Lu ZH, Barton FB, Terrin ML, Charache S, Dover GJ
normal adult blood: all bursts and subcolonies synthesize G gam-
et al. Fetal hemoglobin in sickle cell anemia: determinants of
ma-and A gamma-globin chains. Blood 1980;56:218-26.
response to hydroxyurea. Multicenter Study of Hydroxyurea. Blood
29. Fibach E, Manor D, Treves A, Rachmilewitz EA. Growth of human
normal erythroid progenitors in liquid culture: a comparison with
8. Perrine SP, Greene MF, Faller DV. Delay in the fetal globin switch
colony growth in semisolid culture. Int J Cell Cloning 1991;9:57-64.
in infants of diabetic mothers. N Engl J Med 1985;312:334-8.
30. Myers CD, Katz FE, Joshi G, Millar JL. A cell line secreting stimu-
9. Dover GJ, Brusilow S, Samid D. Increased fetal hemoglobin in
lating factors for CFU-GEMM culture. Blood 1984;64:152-5.
patients receiving sodium 4-phenylbutyrate. N Engl J Med
31. Hale G, Bright S, Chumbley G, Hoang T, Metcalf D, Munro AJ etal. Removal of T cells from bone marrow for transplantation: a
10. Lozzio CB, Lozzio BB. Human chronic myelogenous leukemia
monoclonal antilymphocyte antibody that fixes human comple-
cell-line with positive Philadelphia chromosome. Blood 1975;45:
32. Vaisman B, Konijn AM, Fibach E. Isolation of large and pure sam-
11. Martin P, Papayannopoulou T. HEL cells: a new human erythroleu-
ples of human erythroid precursors at different stages of matura-
kemia cell line with spontaneous and induced globin expression.
tion using immunomagnetic separation of cells from liquid cultures.
12. Komatsu N, Nakauchi H, Miwa A, Ishihara T, Eguchi M, Maroi M
33. Clarke BJ, Housman D. Characterization of an erythroid precursor
et al. Establishment and characterization of a human leukemic
cell of high proliferative capacity in normal human peripheral
cell line with megakaryocytic features: dependency on granulocyte-
blood. Proc Natl Acad Sci USA 1977;74:1105-9.
macrophage colony-stimulating factor, interleukin 3, or erythropoie-
Dalyot N, Fibach E, Rachmilewitz EA, Oppenheim A. Adult and
tin for growth and survival. Cancer Res 1991;51:341-8.
neonatal patterns of human globin gene expression are recapitulat-
13. McHale CM, Winter PC, Lappin TR. Erythroid gene expression is
ed in liquid cultures. Exp Hematol 1992;20:1141-5.
differentially regulated by erythropoietin, haemin and delta-ami-
35. Kollia P, Fibach E, Najjar SM, Schechter AN, Noguchi CT. Modifica-
nolaevulinic acid in UT-7 cells. Br J Haematol 1999;104:829-37. Etions of RNA processing modulate the expression of hemoglobin
14. Rutherford T, Clegg JB, Higgs DR, Jones RW, Thompson J,
genes. Proc Natl Acad Sci USA 1996;93:5693-8.
Weatherall DJ. Embryonic erythroid differentiation in the human
Noguchi CT, Fibach E, Loukopoulos D, Schechter AN.
leukemic cell line K562. Proc Natl Acad Sci USA 1981;78:348-52.
Modulation of globin gene expression in cultured erythroid pre-
15. Wada Y, Noguchi CT. In vitro differential expression of human glo-
cursors derived from normal individuals: transcriptional and post-
bin genes. J Biol Chem 1988;263:12142-6.
transcriptional regulation by hemin. Proc Assoc Am Physicians
16. Gambari R, del Senno L, Barbieri R, Viola L, Tripodi M, Raschella
G et al. Human leukemia K-562 cells: induction of erythroid dif-
37. Dalyot N, Fibach E, Ronchi A, Rachmilewitz EA, Ottolenghi S,
ferentiation by 5-azacytidine. Cell Differ 1984;14:87-97.
Oppenheim A. Erythropoietin triggers a burst of GATA-1 in nor-
mal human erythroid cells differentiating in tissue culture. NucleicAcids Res 1993;21:4031-7.
R. Induction of erythroid differentiation of human K562 cells by 3-
38. Miller JL, Njoroge JM, Gubin AN, Rodgers GP. Prospective identifi-
O-acyl-1,2-O-isopropylidene-D-glucofuranose derivatives. Bioorg
cation of erythroid elements in cultured peripheral blood. Exp
18. Bianchi N, Osti F, Rutigliano C, Corradini FG, Borsetti E, Tomassetti
39. Gelvan D, Fibach E, Meyron-Holtz EG, Konijn AM. Ferritin uptake
by human erythroid precursors is a regulated iron uptake pathway. Blood 1996;88:3200-7.
cells. Br J Haematol 1999;104:258-65.
40. Vaisman B, Fibach E, Konijn AM. Utilization of intracellular ferri-
The DNA-binding drugs mithramycin and chromomycin
are powerful inducers of erythroid differentiation of human K562
tin iron for hemoglobin synthesis in developing human erythroidprecursors. Blood 1997;90:831-8.
. Proc Natl Acad Sci USA 1971;68:1542-6.
41. Meyron-Holtz EG, Vaisman B, Cabantchik ZI, Fibach E, Rouault TA,
Stephenson JR, Axelrad AA, McLeod DL, Shreeve MM. Induction of colonies of hemoglobin-synthesizing cells by erythropoietin inG
Fibach E, Manor D, Oppenheim A, Rachmilewitz EA. Proliferation
Hershko C et al. Regulation of intracellular iron metabolism in
and maturation of human erythroid progenitors in liquid culture.
human erythroid precursors by internalized extracellular ferritin.
21. Wada H, Suda T, Miura Y, Kajü E, Ikemoto S, Yamata Y. Expression
42. Vaisman B, Meyron-Holtz EG, Fibach E, Krichevsky AM, Konijn AM.
Ferritin expression in maturing normal human erythroid precur-sors. Br J Haematol 2000;110:394-401.
43. Pope SH, Fibach E, Sun J, Chin K, Rodgers GP. Two-phase liquid
culture system models normal human adult erythropoiesis at the
ic progenitor cells. Blood 1998;91:2664-71.
molecular level. Eur J Haematol 2000;64:292-303.
of major blood group antigens on human erythroid cells in a twophase liquid culture system. Blood 1990;75:505-11.
22. Malik P, Fisher TC, Barsky LL, Zeng L, Zadi P, Hiti AL et al. An invitro model of human red blood cell production from hematopoiet-
44. Fibach E, Prasanna P, Rodgers GP, Samid D. Enhanced fetal hemo-
sis in culture: II. Heterogeneity in the morphology, time course of
globin production by phenylacetate and 4-phenylbutyrate in eryth-
colony formation, and sedimentation velocities of the colony-
roid precursors derived from normal donors and patients with
forming cells. Am J Hematol 1977;3:29-36.
sickle cell anemia and beta-thalassemia. Blood 1993;82:2203-9.
Eaves CJ, Eaves AC. Erythropoietin (Ep) dose-response curves for
45. Hudgins WR, Fibach E, Safaya S, Rieder RF, Miller AC, Samid D et
three classes of erythroid progenitors in normal human marrow and
al. Transcriptional upregulation of gamma-globin by phenylbuty-
in patients with polycythemia vera. Blood 1978;52:1196-210.
rate and analogous aromatic fatty acids. Biochem Pharmacol
25. Orkin SH, Harosi FI, Leder P. Differentiation in erythroleukemic
cells and their somatic hybrids. Proc Natl Acad Sci USA 1975;72:98-
46. Fibach E, Kollia P, Schechter AN, Noguchi CT, Rodgers GP. Hemin-
induced acceleration of hemoglobin production in immature cul-
Brice M, Stamatoyannopoulos G. Hemoglobin
tured erythroid cells: preferential enhancement of fetal hemoglo-
F synthesis in vitro: evidence for control at the level of primitive
erythroid stem cells. Proc Natl Acad Sci USA 1977;74:2923-7.
47. Fibach E, Schechter AN, Noguchi CT, Rodgers GP. Reducing eryth-
27. Kidoguchi K, Ogawa M, Karam JD. Hemoglobin biosynthesis in
ropoietin in cultures of human erythroid precursors elevates the
individual erythropoietic bursts in culture. Studies of adult periph-
proportion of fetal haemoglobin. Br J Haematol 1994;88:39-45.
eral blood. J Clin Invest 1979;63:804-6.
48. McCaffrey PG, Newsome DA, Fibach E, Yoshida M, Su MS. Induc-
28. Peschle C, Migliaccio G, Covelli A, Lettieri F, Migliaccio AR,
tion of gamma-globin by histone deacetylase inhibitors. Blood
Condorelli M et al. Hemoglobin synthesis in individual bursts from
CULTURE MODELS FOR STUDYING HBF STIMULATION
49. Fibach E. Measurement of total and fetal hemoglobin in cultured
cation to evaluating patients with sickle cell anemia treated with
human erythroid cells by a novel micromethod. Hemoglobin
hydroxyurea. Eur J Haematol 1996;57:17-24.
53. Amoyal I, Goldfarb A, Fibach E. Flow cytometric analysis of hydrox-
50. Wilson JB, Headlee ME, Huisman TH. A new high-performance liq-
yurea effects on fetal hemoglobin production in cultures of beta-
uid chromatographic procedure for the separation and quantita-
thalassemia erythroid precursors. Hemoglobin 2003;27:77-87.
tion of various hemoglobin variants in adults and newborn babies.
54. Fibach E, Burke LP, Schechter AN, Noguchi CT, Rodgers GP.
Hydroxyurea increases fetal hemoglobin in cultured erythroid
51. Huisman TH. Separation of hemoglobins and hemoglobin chains
cells derived from normal individuals and patients with sickle cell
by high-performance liquid chromatography. J Chromatogr 1987;
anemia or beta-thalassemia. Blood 1993;81:1630-5.
55. Hasegawa S, Rodgers GP, Dwyer N, Naguchi CT, Blanchette-Mackie
52. Epstein N, Epstein M, Boulet A, Fibach E, Rodgers GP. Monoclonal
EJ, Uyesaka N et al. Sickling of nucleated erythroid precursors
antibody-based methods for quantitation of hemoglobins: appli-
from patients with sickle cell anemia. Exp Hematol 1998;26:314-9.
TITLE XI: BUSINESS REGULATIONS 110. RETAIL PERMITS CHAPTER 110: RETAIL PERMITS 110.01 Limitation on sale of pharmaceuticals SELL. Furnish, give away, exchange, transfer, deliver, surrender, distribute or supply, whether or not for monetary gain or other WHOLESALER. One whose business is § 110.01 LIMITATION ON SALE OF the selling of goods in gross to retail stores fo
Stepped care bij angst & depressie: Van eerste tot tweede lijn Vrije Universiteit Faculteit der Psychologie en Pedagogiek Afdeling Klinische Psychologie In samenwerking met: Afdeling Huisartsgeneeskunde Afdeling Psychiatrie Drs. Laura Kool Dr. Annemieke van Straten Dr. Harm van Marwijk Prof. Dr. Aartjan Beekman Prof. Dr. Pim Cuijpers INHOUDSOPGAVE 1.1 Achtergrond 1.2
 The use of cell culture procedures
The use of cell culture procedures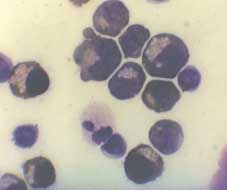
 CULTURE MODELS FOR STUDYING HBF STIMULATION
Figure 2.—Erythroid cells in culture. A) Lar
ge aggregates of erythroid precursors on day 6 of phase II. Unstained cells were photographed
in situ using an inverted microscope. B) Benzidine staining of cells from day 8 phase II cultures. Cells were smeared on a glass slide using
a cytocentrifuge, and stained with benzidine. Mature erythroid Hb-containing cells which are benzidine positive (B+) and early erythroid pre-cursors which are benzidine negative (B-) are seen. C) Benzidine/Giemsa staining of cells from day 10 cultures. Cells were smeared as in Bfollowed by staining with Giemsa. Both B+ erythroid cells and B- myeloid and lymphoid cells are seen. D) Cells harvested on day 12 of theculture.
CULTURE MODELS FOR STUDYING HBF STIMULATION
Figure 2.—Erythroid cells in culture. A) Lar
ge aggregates of erythroid precursors on day 6 of phase II. Unstained cells were photographed
in situ using an inverted microscope. B) Benzidine staining of cells from day 8 phase II cultures. Cells were smeared on a glass slide using
a cytocentrifuge, and stained with benzidine. Mature erythroid Hb-containing cells which are benzidine positive (B+) and early erythroid pre-cursors which are benzidine negative (B-) are seen. C) Benzidine/Giemsa staining of cells from day 10 cultures. Cells were smeared as in Bfollowed by staining with Giemsa. Both B+ erythroid cells and B- myeloid and lymphoid cells are seen. D) Cells harvested on day 12 of theculture.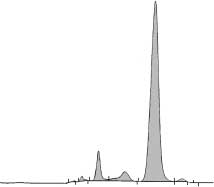 CULTURE MODELS FOR STUDYING HBF STIMULATION
tains progenitors at various developmental stages.33
Good results can be obtained with cells derived fromother sources, including CD34+ cells purified by immu-
no-magnetic bead technologies, but, in these cases
some modifications of the procedure are required.
CULTURE MODELS FOR STUDYING HBF STIMULATION
tains progenitors at various developmental stages.33
Good results can be obtained with cells derived fromother sources, including CD34+ cells purified by immu-
no-magnetic bead technologies, but, in these cases
some modifications of the procedure are required.
 CULTURE MODELS FOR STUDYING HBF STIMULATION
M T Glycophorin A
CULTURE MODELS FOR STUDYING HBF STIMULATION
M T Glycophorin A