He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Untitled
TEJIDO NERVIOSO. MSc. Belén Z. Iglesias Ramírez Dr. Andrés Dovale Borjas
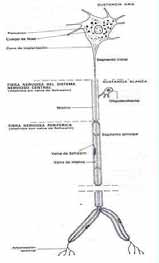
El tejido nervioso, al igual que los demás tejidos básicos, está compuesto por células, sustancia intercelular y líquido tisular. Los elementos celulares que lo integran son: neuronas y neuroglias. Las neuronas se distinguen por su aspecto morfológico, presentan un soma o cuerpo y prolongaciones citoplasmáticas que se denominan axón y dendrita
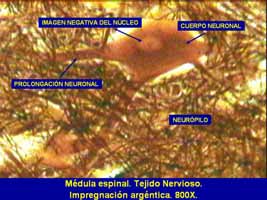
A B Fig. 1. A. Esquema de una neurona multipolar . B . Beurona multipolar de la médula espinal. Impregnación argéntica. La función de las neuronas está basada en el desarrollo de dos propiedades fundamentales del protoplasma, excitabilidad y conductividad. Las neuronas son las encargadas de recibir los estímulos del medio, transformarlos en excitaciones nerviosas y trasmitirlos a los centros nerviosos, en los que se organizan para dar una respuesta. Por su parte, las neuroglias cumplen funciones nutritivas, aislantes, de sostén y defensa. Las prolongaciones largas del cuerpo de la neurona (axón) constituyen la parte fundamental de las fibras nerviosas, las que se entremezclan en la mayor parte de los órganos del sistema nervioso, con dendritas y con prolongaciones de las neuroglias. Este conjunto de fibras entrecruzadas constituyen el neurópilo. El tejido nervioso es el componente fundamental de una serie de órganos, cuyo conjunto se denomina sistema nervioso. El sistema nervioso está compuesto por el sistema nervioso central (SNC), que incluye el encéfalo y la médula espinal, y el sistema nervioso periférico (SNP) formado por los nervios craneales, los nervios raquídeos, los ganglios nerviosos y las terminaciones nerviosas. Neuronas Están constituidas por un cuerpo celular o soma y las prolongaciones, algunas de más de un metro de largo. Entre dichas prolongaciones se distinguen el: axón
(transmisor del impulso nervioso), que es uno solo para cada neurona; y las dendritas (receptoras del impulso nervioso), generalmente múltiples. El tamaño del cuerpo o soma de las neuronas varía desde muy pequeño, de 4 a 6 µm, en las llamadas células granulosas o granos del cerebelo, hasta de 150 µm en las células piramidales gigantes de Betz del área motora de la corteza cerebral.
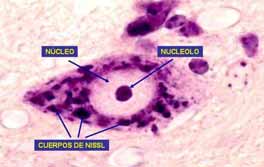
A B Fig. 2. A. Esquema del cuerpo de una neurona multipolar. B, Microfotografía de una neurona multipolar. Cresil violeta. Se observa el nucleolo y los cuerpos de Nissl. La forma de las neuronas también es variada, debido principalmente al número y la disposición de sus prolongaciones. Las neuronas pueden ser estrelladas, fusiformes, piramidales, esféricas, etc. DISTRIBUCIÓN. En el SNC los cuerpos neuronales se agrupan en la corteza cerebral, corteza cerebelosa y en los núcleos grises. Estas zonas en estado fresco presentan un color grisáceo dada la abundancia de cuerpos neuronales y poca presencia de fibras nerviosas mielínicas, a estas zonas se le denomina sustancia gris. En la sustancia gris, además de los somas neuronales y sus prolongaciones, se encuentran gran número de células de neuroglia y capilares sanguíneos. Las zonas del SNC donde predominan las fibras nerviosas mielínicas (axones revestidos de mielina) se les denomina sustancia blanca, ya que por el alto contenido en lípidos de la mielina estas zonas presentan color blanco. En el SNP los cuerpos neuronales se agrupan en los ganglios nerviosos del Sistema Nervioso Autónomo. CLASIFICACIÓN MORFOLÓGICA DE LAS NEURONAS De acuerdo al número de prolongaciones dendriticas las neuronas se clasifican en:
Fig. 3 Clasificación de las neuronas según el número de dendritas. Unipolares: Son las que poseen una sola prolongación que parte del cuerpo neuronal. Las neuronas unipolares son muy raras en el humano, pueden verse durante el desarrollo embrionario (neuroblastos unipolares) y en la retina las células amacrinas. Pseudounipolares: Las que se encuentran en los ganglios sensitivos de la raíz dorsal de los nervios espinales y en los ganglios sensitivos de varios nervios craneales. Estas células derivan de neuroblastos bipolares y durante su desarrollo, las prolongaciones se fusionan en su parte proximal por lo que la neurona queda con una sólo prolongación que se bifurca a cierta distancia del cuerpo neuronal. Los procesos resultantes, por su estructura y su capacidad para conducir los impulsos nerviosos, son axones por lo que las neuronas pseudounipolares no poseen dendritas. (fig. 9.2). Las neuronas bipolares poseen una dendrita y un axón que se localizan en polos opuestos de la célula. La dendrita puede estar o no ramificada y el axón puede ser corto o largo. Este tipo de neuronas se puede encontrar en la retina y en los ganglios vestibulares y cocleares del oído interno (fig. 9.2). Por la forma de huso del soma celular estas células son fusiformes. Las neuronas multipolares son las más abundantes del sistema nervioso; en ellas el soma celular presenta más de una prolongación dendrítica. Presentan un solo axón (fig. 9.2) El soma de estas neuronas puede ser estrellado, piramidal, piriforme, etc.
Fig. 4. Diferentes formas de neuronas multipolares El histólogo italiano Camilo Golgi, clasificó las neuronas según la longitud del axón en:
Las neuronas Golgi tipo I poseen axones largos que salen de la región donde se encuentra el soma celular y terminan lejos de su origen, en otra parte del sistema nervioso o en otro tejido, tal como la piel o los músculos. Las neuronas tipo II poseen axones cortos que se ramifican localmente en la región donde se sitúa el soma neuronal (figura 9.3). CARACTERÏSTICAS MORFOFUNCIONALES DE LAS NEURONAS: CUERPO O SOMA NEURONAL. El cuerpo de la neurona constituye el centro trófico o nutricio de la célula y proporciona una gran área de superficie de membrana para recibir los impulsos nerviosos. Los dos componentes del soma neuronal son el núcleo y el pericarion. El pericarion es el citoplasma neuronal que rodea al núcleo (peri, alrededor; cario, núcleo). La neurona por lo general tiene una estructura típica de célula productora de proteínas. Esto se hace más evidente en las grandes células multipolares, NÚCLEO. El núcleo de las neuronas es generalmente voluminoso (6-10 µm), esférico y de cromatina laxa. Poseen uno o dos nucléolos prominentes que se destacan en la matriz nuclear. La envoltura nuclear de las neuronas presenta numerosos poros nucleares y adosada a su cara interna se encuentra la cromatina periférica.
Fig. 5. Se observa el cuerpo de una neurona seudounipolar del ganglio craneoespinal. En el núcleo se observa el nucléolo y en el citoplasma pigmento de lipofucsina. PERICARION.
El pericarion está delimitado por la membrana celular y rodeando al núcleo. Del pericarion parten los procesos celulares: dendritas y axones. En el mismo es donde se realizan las funciones metabólicas y biosintéticas esenciales. Los procesos glucolíticos, incluidos el ciclo de Krebs mitocondrial, son muy activos; las células consumen más de 100 g. de glucosa en 24 h. La membrana celular es de gran importancia, pues de su actividad dependen el origen y la propagación de los impulsos nerviosos. En la composición química de las membranas
plasmáticas de las neuronas se incluye la presencia de glucoproteínas en la porción externa de la membrana. con la presencia de ácido siálico, cuya carga negativa podría relacionarse con la fijación del Ca++ y Na+, y de enzimas para permitir la entrada activa de potasio y la salida de sodio. Neuroplasma, es la parte amorfa de citoplasma, en el se observan; neurofibrillas, sustancia cromófila o cuerpos de Nissl, mitocondrias, aparato de Golgi e inclusiones. Las neurofibrillas se hacen evidentes con las técnicas de impregnación con sales de plata y en las coloraciones vitales con azul de metileno, y se presentan en forma de malla en todo el pericarion y se extiende a las prolongaciones. Al M/E las neurofibrillas se observan formadas por haces de microtúbulos y microfilamentos que forman el citoesqueleto neuronal y participan en el transporte de sustancias y organitos celulares hacia las prolongaciones y de ellas al cuerpo celular. Los cuerpos de Nissl son gránulos basófilos abundantes en los cuerpos neuronales. Los cuerpos de Nissl se encuentran diseminados en el neuroplasma del pericarion y dendritas de mayor diámetro, pero están ausentes en el cono axónico y el axón. Al M/E se corresponden con la intensa presencia de ribosomas adosados a membranas de retículo endoplasmático rugoso; también los ribosomas pueden encontrarse libres y formando polirribosomas. Todo ello indica la intensa actividad de síntesis proteica en las neuronas. Como corresponde a células metabólicamente muy activas, las neuronas contienen muchas mitocondrias, localizadas en el pericarion. También se encuentran mitocondrias en las dendritas y en el axón. El aparato de Golgi está muy desarrollado en las células nerviosas, y al M/O utilizando técnicas de plata se observa como una malla reticular alrededor de núcleo. Al M/E se visualiza como varios grupos de sacos de paredes aplanadas (dictiosomas). Se observan además los otros componentes del Golgi como son; vesículas pequeñas, y gránulos secretorios, muy desarrollados en las neuronas neurosecretoras de los núcleos hipotalámicos. El pericarion neuronal también posee lisosomas, que aparecen como cuerpos densos asociados al aparato de Golgi. Como producto final de la digestión lisosomal en la neurona se forman cuerpos residuales (gránulos de lipofucsina), cuyo número aumenta con la edad del individuo. En algunas enfermedades metabólicas el aumento de los cuerpos residuales es de tal magnitud que afecta el funcionamiento de las neuronas. Existen en las neuronas varios tipos de inclusiones: Lipofucsina, de color amarillento, que se incrementa con la edad y representan residuos insolubles de la actividad lisosomal (cuerpos residuales). Al M/E se visualiza como un cuerpo heterogéneo, rodeado por una membrana donde alternan partículas densas y lípidos formando goticas o membranas enrolladas sobre si mismas (figura de mielina). Melanina, se aprecia fundamentalmente en la sustancia negra del cerebro medio, en el locus niger y en otras regiones. Su significación biológica no está bien esclarecida. Glucógeno, puede encontrarse en las células nerviosas embrionarias. No es frecuente su localización en neuronas adultas. Gránulos que contienen hierro, se encuentra en la sustancia negra, y en el globus pallidus. Lípidos, se pueden observar en neuronas adultas algunas gotas pequeñas de lípido como expresión de reserva metabólica o de alteración del funcionamiento celular. PROLONGACIONES.
Las prolongaciones del cuerpo neuronal son las dendritas y el axón.
Las dendritas son generalmente múltiples, cortas y ramificadas. En su origen son más anchas que el axón y se van adelgazando a medida que se ramifican alejándose del cuerpo neuronal. Contienen la mayoría de los organitos típicos del pericarion (cuerpos de Nissl, mitocondrias y neurofibrillas). Al M/E se observan RER, ribosomas libres y componentes del complejo de Golgi, solo en la porción proximal, es decir, cerca del pericarion. Las dendritas se ramifican dando lugar a ramas de menor diámetro; a medida que esto ocurre, el RER y los otros organitos son menos frecuentes. Los microtúbulos y microfilamentos llegan hasta los procesos más finos. Un carácter sumamente importante en las dendritas es la presencia, en su superficie, de múltiples contactos sinápticos. En estos puntos de contacto sináptico las dendritas muestran pequeñas proyecciones denominadas espinas dendríticas.
En la mayoría de las neuronas las dendritas son cortas, ramificándose cerca del cuerpo celular. Su número, longitud y terminación varían en extremo y no dependen del tamaño del pericarion. Las dendritas, a través de sus sinapsis reciben impulsos nerviosos de otras neuronas.
El axón o cilindroeje, es una prolongación única, de diámetro variable y de hasta 100 cm de longitud. El axón conduce al impulso desde el soma hacia otras neuronas, músculos o glándulas. El axón puede recibir también estímulos de otras neuronas, con lo que se modifica su función. Difiere considerablemente de las dendritas; mientras hay usualmente varias dendritas, existe sólo un axón en cada neurona. Esta prolongación se origina en una región de forma cónica en el cuerpo celular que no posee cuerpos de Nissl y que se denomina cono axónico. El axón es generalmente más delgado, su diámetro se mantiene constante hasta la arborización terminal y es más largo que las dendritas de las mismas neuronas.
A lo largo de su curso el axón puede emitir ramas colaterales; no obstante, su arborización principal ocurre en su terminación y está compuesta por ramas primarias, secundarias y yemas, las cuales son variables en número, forma y distribución. A menudo estas ramas forman mallas que rodean a neuronas relacionadas, o se pliegan alrededor de las dendritas de otras neuronas. Al extremo ramificado del axón se le denomina telodendrón y a la terminación abultada del extremo de cada ramificación se le denomina botón terminal o botón sináptico. Existen otras terminaciones del axón como son las placas motoras musculares, terminaciones en cesta, terminaciones de receptores especiales o generales, terminaciones anuloespirales del huso neuromuscular, etc., que varían considerablemente la forma de la terminación axónica, por lo que se dice que la misma es bastante variable desde el punto de vista morfológico.
En el cono axónico no se observan cuerpos de Nissl, detalle que sirve para diferenciar el axón de los demás procesos dendríticos de la neuronas. Al M/E, en el axón se pueden observar mitocondrias, vesículas de superficie lisa, microfilamentos y microtúbulos. En las neuronas secretoras se pueden encontrar gránulos secretorios. Las mitocondrias, microfilamentos y microtúbulos se disponen longitudinalmente, es decir siguiendo el eje mayor del axón. Una característica del axón es que, al igual que el cono axónico, no presenta gránulos de Nissl ni RER, lo que unido a la presencia de vesículas conteniendo secreción, permite establecer la diferenciación entre una dendrita y el axón al M/E.
El axón trasmite normalmente excitaciones nerviosas que se originan en el cono axónico. Estas excitaciones son trasmitidas a través de las sinapsis a otras neuronas o a células efectoras, tales como las fibras musculares o las células glandulares. El contacto celular
entre axones y dendritas, o axones y cuerpos celulares se denomina sinapsis.
NEUROGLIAS.
Son células cuya función es el sostén metabólico, mecánico y la protección de las neuronas. Las neuroglias se caracterizan por ser mucho más numerosas, puede haber hasta 10 veces más células de neuroglia que neuronas en el sistema nervioso, y, generalmente, de menor tamaño que las neuronas. En los cortes histológicos de rutina sólo se visualizan sus núcleos, ubicados entre los cuerpos neuronales y entre los haces de fibras. Las Neuroglias se presentan tanto en el Sistema Nervioso Central como en el Sistema Nervioso Periférico- NEUROGLIA CENTRAL.
En el Sistema Nervioso Central las glías se clasifican en Macroglías, Microglías y Células Ependimarias. La macroglía incluye los astrocitos y la oligodendroglía. Astrocitos Los astrocitos son las más grandes de las células de neuroglia, y existen en dos tipos diferentes: astrocitos protoplásmicos en la sustancia gris del SNC, y astrocitos fibrosos que se encuentran principalmente en la sustancia blanca del sistema nervioso central. Los astrocitos son células estrelladas en las que su cuerpo celular da lugar a prolongaciones de longitud y grosor variables que se ramifican entre las neuronas. Los astrocitos protoplasmáticos poseen núcleos ovales y vesiculares. Sus prolongaciones son más cortas, gruesas y ramificadas que en los astrocitos fibrosos. Algunas de sus prolongaciones terminan en expansiones laminares alrededor de la membrana basal de los capilares sanguíneos formando los llamados pies vasculares, otras sobre el cuerpo o las dendritas de las neuronas y aún otras forman parte de la membrana piaglial que recubre al SNC. Los astrocitos fibrosos poseen en su citoplasma sólo unos cuantos organitos, ribosomas libres y glucógeno. Las prolongaciones de esas células, de 20 a 40, son finas, largas, rectas y, sobre todo, no ramificadas, dándole a la célula su aspecto típico de araña en las impregnaciones argénticas. Al igual que los protoplasmáticos, sus prolongaciones se aplican a los capilares sanguíneos, conformando pies vasculares, Los astrocitos funcionan como depredadores de iones y residuos del metabolismo neuronal, como iones K+, glutamato y ácido gamma-aminobutírico, que se acumulan en el microambiente de las neuronas. También contribuyen al metabolismo energético dentro de la corteza cerebral al descargar glucosa, a partir de su glucógeno almacenado. Los pies vasculares intervienen en el metabolismo de la neurona, de forma tal que los productos tóxicos, medicamentosos o nutritivos, que se encuentran en la sangre, antes de llegar a la neurona son metabolizados por los astrocitos que forman parte de la barrera hematoencefálica. En las zonas lesionadas los astrocitos se acumulan para formar tejido de cicatrización. Oligodendroglia. Se parecen a los astrocitos, pero son más pequeños y contienen menos prolongaciones con ramificaciones escasas y núcleo pequeño, esférico y de cromatina más densa. Son las células de neuroglia que toman las tinciones más oscuras, están localizadas en las sustancias tanto gris como blanca del sistema nervioso central. Su citoplasma denso contiene un RER abundante, muchos ribosomas libres y mitocondrias, y un complejo de Golgi definido. Contienen también microtúbulos, sobre todo en la zona perinuclear y en las prolongaciones celulares. Se disponen entre haces de axones (interfasciculares) y alrededor de las neuronas. Los oligodendrocitos son los encargados de elaborar y conservar la mielina sobre los axones del SNC. Al producir mielina, los oligodendrocitos funcionan de manera semejante a las células de Schwann del SNP. Un sólo oligodendrocito puede envolver a varios axones, en tanto que la célula de Schwann envuelve sólo a un axón. A B Fig. 6 . A. Se muestran los oligodendrocitos. B. Formación de mielina por los oligodendrocitos Microglia. Son los macrófagos del sistema nervioso. Diseminadas por todo el SNC, las células de microglia son pequeñas y de tinción oscura. Su citoplasma es escaso; un núcleo oval y prolongaciones cortas e irregulares. El cuerpo celular y sus prolongaciones están también tachonados de espinas. Estas células funcionan como fagocitos para eliminar los desechos y las estructuras lesionadas en el sistema nervioso central. A diferencia de las otras células de neuroglia, que se derivan del tubo neural, las células de microglia se originan en la médula ósea y son parte del Sistema de Macrófagos. Células ependimarias. Son células cilíndricas a cúbicas bajas, con núcleo esférico y central, que se disponen formando membranas de una sola célula de grosor que revisten los ventrículos cerebrales y el conducto central de la médula espinal. Se derivan del neuroepitelio embrionario del sistema nervioso en desarrollo. Su citoplasma contiene abundantes mitocondrias y haces de filamentos intermedios. Algunas presentan microvellosidades apicales y en algunas regiones son ciliadas, y por este motivo facilitan el movimiento del líquido cefalorraquídeo. Las prolongaciones que salen del cuerpo celular llegan a la superficie del cerebro en el embrión, pero en el adulto se encuentran reducidas y terminan en las células cercanas. Su función principal es la formación, intercambio y circulación del líquido cefalorraquídeo. Durante el desarrollo embrionario participa en la modelación de la citoarquitectura del SNC. NEUROGLIA PERIFÉRICA La Neuroglia periférica está constituida por : las células de Schwann, las células satélites y las células de Mueller. Estas últimas se estudiaran en la retina. Células de Schwann. Las células de Schwann son células aplanadas cuyo citoplasma contiene un núcleo aplanado, un aparato de Golgi pequeño, y unas cuantas mitocondrias. El resto del citoplasma de la célula de Schwann con el núcleo, queda rodeando la vaina de mielina y se le denomina neurilema o vaina de Schwann.: La microscopio electrónica ha revelado que la mielina es el plasmalema de las células de Schwann organizada en una vaina que se envuelve múltiples veces alrededor del axón. En la vaina de mielina se encuentran interrupciones a intervalos regulares a toda la Iongitud del axón, que se denominan nodos de Ranvier, sitios en los que queda expuesto el axón. La porción externa de las células de Schwann está cubierta por una lámina basal que se sumerge en los nodos de Ranvier. Después de una lesión nerviosa, el nervio en regeneración se encuentra guiado por la lámina basal. Las zonas del axón cubiertas por las laminillas concéntricas de mielina y la célula de Schwann única que produjo la mielina se denominan segmentos internodales, y su longitud varía entre 200 y 1000 µm. Una célula de Schwann, puede mielinizar sólo un internodo de un sólo axón, en tanto que las células de oligodendroglia, pueden mielinizar a los internodos de varios axones. Aunque una célula de Schwann puede mielinizar sólo a un axón, varios axones amielínicos pueden estar envueltos por una sola célula de Schwann. Células capsulares. Las células satélites o capsulares constituyen células de sostén que rodean los cuerpos neuronales. En las neuronas pseudounipolares de los ganglios de la raíz posterior de los nervios espinales es donde mejor se distinguen dispuestas a modo de cápsula celular alrededor de los cuerpos neuronales, de ahí su nombre de células capsulares. Estas glías periféricas son de pequeño tamaño y tienen escaso citoplasma. Las glías, en general, están involucradas en las siguientes funciones:
• Ciertas glías producen las vainas de mielina: en el sistema nervioso central (SNC),
son las oligodendroglias, y en el sistema nervioso periférico (SNP), las células de Schwann.
• Muestran propiedades fagocíticas: microglias.
• Durante el desarrollo embrionario las fibras gliales guían, tanto la migración de las
neuronas como el crecimiento de sus prolongaciones.
• Los astrocitos forman parte de la barrera hematoencefálica.
• En intercambios macromoleculares, metabólicos y nutricionales, que pueden ocurrir
por la vía de interacciones de los constituyentes de superficie o por la vía de transferencia de macromoléculas de una célula a otra. Existen uniones espaciadas entre las glías y las neuronas, de manera que las glías y las neuronas operan como unidades acopladas metabólicamente.
• Aunque existen uniones comunicantes o de hendidura entre las células de
neuroglia, éstas no reaccionan con los impulsos nerviosos ni los propagan.
FORMACIÓN DE LA MIELINA.
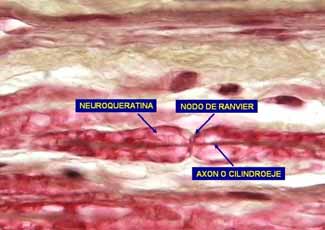
Al M/O la vaina de mielina aparece como un tubo alongado que se interrumpe a intervalos regulares en los denominados nodos de Ranvier. Los segmentos de mielina entre nudos consecutivos se denominan internodos. En el SNC cada segmento de mielina está formado por una prolongación citoplasmática de una oligodendroglia que se dirige hacia el axón y da varias vueltas alrededor del mismo, enrollándolo con una vaina formada por capas yuxtapuestas de la membrana celular, la vaina de mielina. No está claro el mecanismo de la mielinización, proceso por medio del cual la célula de Schwann (o el oligodendrocito) envuelve su membrana de manera concéntrica alrededor del axón para formar la vaina de mielina. Se cree que se inicia conforme la célula de Schwann va envolviendo al axón y, de alguna manera, envuelve también su membrana alrededor de éste. La envoltura puede proseguir por más de 50 vueltas. Conforme la membrana se enrolla alrededor del axón, produce una serie de líneas densas amplias alternantes con líneas menos densas y más estrechas a intervalos de 12 nm. La microscopía electrónica de alta resolución ha revelado un pequeño espacio dentro de la línea interperiódica entre cada capa espiral de la vaina de mielina, que se denomina espacio interperiódico. Se considera que este espacio ofrece acceso a las pequeñas moléculas para que lleguen al axón. La región de la línea interperiódica que está en contacto íntimo con el axón se conoce como mesoaxón interno, en tanto que su superficie más externa, en contacto con la célula de Schwann es el mesaron externo.
A B Fig. 7. Nervio periférico. A microfotografía electrónica. B microscopía óptica. La microscopía de luz ha revelado diversos surcos oblicuos de forma cónica en la vaina de mielina de cada segmento internodal que se denominan hendiduras de Schmidt-Lantermann. Se demostró que estas hendiduras, vistas con el microscopio electrónico, eran citoplasma de las células de Schwann atrapado dentro de las vainas de mielina. Un axón adquiere su vaina mielínica al final del segmento inicial, es decir, un poco después del cono axónico, y continúa hasta cerca de la terminación axónica. Las porciones desnudas de los nodos de Ranvier son regiones altamente especializadas, responsables de la capacidad autorregenerativa del potencial de acción conducido. Durante el paso de un potencial de acción cambia la conductancia de la membrana entre los nudos, de manera que la onda de despolarización "salta" de nodo a nodo y forma la
conducción "saltatoria", que es mucho más rápida que en las fibras amielínicas. Las fibras amielínicas en el SNC están también revestidas por oligodendroglias, pero no poseen mielina. SINAPSIS.
La sinapsis se define como el contacto de los extremos finales (botones terminales) de los axones neuronales con una porción de membrana de otra célula. Pueden existir tres tipos de contacto:
1. Sinapsis neuroneuronal, cuando el contacto se establece entre dos neuronas. 2. Sinapsis neuromuscular, cuando el contacto se establece entre el botón
sináptico y la superficie de una célula muscular.
3. Sinapsis neuroepitelial, cuando el contacto se establece entre la neurona y una
También algunos autores consideran las terminaciones nerviosas sensoriales como tipo especializado de sinapsis, aún cuando la fibra nerviosa contacta con células o estructuras derivadas de las mismas que no reúnen características de células nerviosas, musculares o epiteliales típicas.
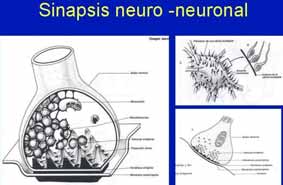
Fig 8. Sinapsis neuro-neuronal Una sinapsis neuroneuronal puede definirse como el contacto entre un botón terminal o sináptico y una porción de membrana de otras células nerviosas donde, mediante una serie de especializaciones morfológicas, ocurre la liberación de un agente químico neurotransmisor del axón que influye con la conductancia de la célula receptora. Las sinapsis neuroneuronales se clasifican de acuerdo con la zona celular con la que el botón sináptico establece el contacto:
• Sinapsis axosomática, cuando el bulbo axónico establece sinapsis con el cuerpo de
• Sinapsis axodendrítica, cuando el botón terminal establece contacto con las
dendritas de otra neurona. Generalmente con las espinas dendríticas.
• Sinapsis axoaxónicas, cuando el botón terminal axónico contacta con otro axón.
Una sinapsis típica consta de un elemento presináptico y otro postsináptico en estrecha asociación mediante regiones especializadas de sus membranas plasmáticas, separados ambos solamente por una estrecha hendidura extracelular de 20-30 nm, la hendidura sináptica.
El lado presináptico presenta un acúmulo de material electrondenso en
forma de placa adosado a la cara interna de la membrana celular, este material aparece delimitando espacios cilíndricos, hexagonales, de citoplasma claro, a manera de túneles que permiten el paso de las vesículas sinápticas hacia la membrana celular (rejilla presináptica). Filamentos de actina se asocian a esta placa densa y se ramifican por todo el botón sináptico. Las vesículas sinápticas de centro claro, o denso, que miden de 40-60 nm. de diámetro, contienen al neurotransmisor. En este lado de la sinapsis podemos encontrar también mitocondrias, REL y vesículas de cubierta. Las vesículas del REL y las de cubierta tienen relación con el recambio de neurosecreción y de las membranas celulares a partir de la pérdida del equilibrio que se presenta con la liberación del contenido de las vesículas sinápticas en la hendidura sináptica. Durante el proceso de secreción nos solo se libera la neurosecreción en la hendidura sináptica, sino que además la membrana de la vesícula queda incorporada a la membrana celular, esto último produce un exceso de membrana en la superficie celular que es removido por la formación de vesículas de cubierta. Durante este proceso se plantea que también se recupera el neurotransmisor.
• La hendidura sináptica es el espacio intercelular que existe entre las estructuras pre
y postsinápticas. Es un espacio de 20 a 30 nm (0.2 nm en las sinapsis eléctricas) que presenta material electrondenso en forma de filamentos que parecen unir a la membrana presináptica con la postsináptica y que parecen corresponder con glucoproteínas transmembrana.
• En el lado postsináptico no se pueden encontrar vesículas sinápticas y si RER,
microtúbulos y microfilamentos. En las sinapsis denominadas asimétricas, se encuentra un engrosamiento mayor que la rejilla presináptica o engrosamiento presináptico. Pero el éste puede faltar en algunos tipos de sinapsis, denominadas asimétricas por este motivo.
Cuando el potencial de acción llega a la membrana presináptica, inicia la apertura de canales de Ca++, lo que permite que entren éstos iones. La entrada de Ca++ hace que las vesículas sinápticas se fusionen con la membrana presináptica y vacíen su neurotransmisor en la hendidura sináptica por medio de exocitosis. El exceso de membrana se recaptura por medio de endocitosis mediada por clatrina. Las vesículas de cubierta formadas se fusionan con el REL, sitio en el que se recicla de manera continua la nueva membrana. El neurotransmisor se difunde a través de la hendidura sináptica hasta receptores de canales iónicos de compuerta situados sobre la membrana postsináptica. La fijación del neurotransmisor a éstos receptores inicia la apertura de los canales iónicos que permite el paso de ciertos iones, lo que altera la permeabilidad de la membrana postsináptica e invierte su potencial de membrana. Es importante percatarse que los neurotransmisores no producen los sucesos a nivel de la membrana postsináptica; sólo la activan. Cuando el estímulo a nivel de una sinapsis da por resultado despolarización de la membrana postsináptica hasta un valor umbral que inicia un potencial de acción, se denomina potencial postsináptico excitatorio. El estímulo a nivel de la sinapsis que da por resultado conservación del potencial de la membrana o aumento de la hiperpolarización de ésta se denomina potencial postsináptico inhibitorio.
El espesor y las densidades relativas de las membranas presináptica y postsináptica acopladas y la amplitud de la hendidura sináptica se correlacionan, generalmente, con la naturaleza de la reacción. Una densidad postsináptica gruesa y una hendidura sináptica de 30 nm se denominan, en conjunto, sinapsis asimétrica, que suele ser el sitio de las reacciones excitatorias. La densidad postsináptica delgada con un hendidura sináptica de 20 nm constituyen una sinapsis simétrica, que suele ser el sitio de las reacciones inhibitorias. La excitación nerviosa se propaga a lo largo de la membrana plasmática por el potencial de acción. Este consiste en una despolarización súbita de la membrana acompañada del aumento de la permeabilidad al Na+. El potencial cambia de -70 mV. a más de +50 mV. El potencial de acción tiene un umbral de activación, es una respuesta del tipo todo o nada, no decrece y presenta un periodo refractario durante el cual no puede reaccionar a otro estímulo. SINAPSIS ELÉCTRICAS.
Aunque las sinapsis eléctricas son poco frecuentes en los mamíferos, se encuentran en el tallo cerebral, la retina y la corteza cerebral. Las sinapsis eléctricas suelen estar representadas por uniones comunicantes o de hendidura (nexos) que permiten el paso libre de iones desde una célula hacia otra. Cuando sucede así entre neuronas, el resultado será un flujo de corriente. La transmisión de impulsos es mucho más rápida a través de las sinapsis eléctricas que a través de las químicas. FIBRAS NERVIOSAS.
Las fibras nerviosas son estructuras largas y delgadas, especializadas en la conducción de los impulsos nerviosos, están constituidas por un axón y una vaina producida por células gliales que se disponen a continuación una de otra a lo largo de todo el trayecto del axón. En el SNP existen fibras nerviosas amielínicas y mielínicas, dependiendo de si las células gliales que envuelven al axón producen o no la vaina de mielina. Las fibras nerviosas amielínicas están constituidas por varios axones que se empotran en canales formados por invaginaciones de la membrana celular de las células de Schwann que forman la vaina de la fibra nerviosa (figura 9.8). En secciones histológicas transversales de una fibra nerviosa amielínica se pueden observar hasta 12 axones o más empotrados en la célula de Schwann. En las fibras nerviosas mielínicas, los axones son rodeados por las células de Schwann, y la lengüeta citoplasmática de un lado, va formando láminas que al compactarse alrededor del axón forman la estructura periódica ya descrita anteriormente. CORRELACION HISTOFISIOLOGICA EN EL TEJIDO NERVIOSO.
Las neuronas realizan funciones tróficas y metabólicas comunes a otras células del organismo; sin embargo, están altamente especializadas en dos propiedades, la excitabilidad y la conductividad. Las neuronas y otras células se encuentran polarizadas desde el punto de vista eléctrico con un potencial en reposo de cerca de -70 mV (el interior es menos positivo que el exterior) a través de la membrana plasmática. En la mayor parte de las células el potencial es, por lo general, constante. Sin embargo, en neuronas y células musculares el potencial de la membrana puede experimentar cambios controlados, lo que vuelve a estas células capaces de conducir una señal eléctrica. Este potencial se origina a causa de las diferencias en las concentraciones de iones dentro y fuera de la célula. En las células de mamíferos, la concentración de iones K+ es mucho más elevada dentro de la célula que fuera de ella, en tanto que la concentración de iones Na+ y CI- es mucho más elevada fuera de las células que en su interior. Aunque la conservación del potencial en reposo depende, principalmente, de los canales de fuga de K+, las bombas de Na+ y K+ de la membrana plasmática ayudan al bombear activamente iones Na+ hacia el exterior de la célula e iones K+ hacia el interior de ésta. Por cada tres iones Na+ que se bombean hacia el exterior, entran en la célula dos iones K+. Una neurona recibe e integra múltiples estimulaciones a través de las sinapsis, las recibidas por las dendritas se suman a las recibidas en el soma de modo que el potencial eléctrico de la membrana celular acaba por exceder al umbral y origina un impulso nervioso en la zona del cono axónico. Los impulsos nerviosos son señales eléctricas generadas por las zonas desencadenantes de espigas (conos axónicos) de una neurona como resultado de despolarización de la membrana, que se conducen a lo largo del axón hasta su terminación. La transmisión de impulsos desde las terminaciones de una neurona hacia otra neurona, una célula muscular o una glándula se produce a nivel de las sinapsis. La estimulación de una neurona produce apertura de canales de Na+ de compuerta de voltaje en una pequeña región de la membrana, lo que produce entrada de Na+ en la célula por ese sitio. El exceso local de iones Na+ en el interior de la célula produce inversión del potencial en reposo (es decir, el interior se vuelve positivo en relación con el exterior), y se dice que la membrana está despolarizada. Esto provoca el cierre de los canales de Na+ durante 1 a 2 ms, lo que se conoce como periodo refractario, en el cual los canales cerrados están inactivos y no pueden abrirse. Durante este periodo se abren los canales de K+ de compuerta de voltaje, y permiten la salida de estos iones hacia el líquido extracelular con lo que se restaura el potencial de la membrana en reposo; sin embargo, puede ocurrir un periodo breve de hiperpolarización. Una vez restaurado el potencial en reposo, los canales de K+ de compuerta de voltaje se cierran y termina el periodo refractario. El ciclo de despolarización e hiperpolarización de la membrana y de retorno al potencial de la membrana en reposo se denomina potencial de acción, reacción del todo o nada que puede ocurrir a ritmos de hasta 1 000 impulsos/segundo. La despolarización de la membrana que ocurre al abrirse los canales de Na+ de compuerta de voltaje en un punto de un axón se extiende, de manera pasiva, a corta distancia y desencadena la apertura de los canales adyacentes, lo que da por resultado generación de otro potencial de
acción. De esta manera la onda de despolarización, o impulso nervioso, se conduce a lo largo del axón. In vivo un impulso se conduce sólo en una dirección, desde el sitio de la despolarización inicial hasta la terminal del axón. La inactivación de los canales Na+ cerrados durante los periodos refractarios impide la propagación retrógrada de la onda de despolarización. El impulso nervioso es conducido a lo largo de las fibras nerviosas, hasta sus terminaciones, allí a través de las sinapsis estimula a otra célula nerviosa o a otra célula de tipo efector. Las fibras nerviosas son mielínicas y amielínicas. La vaina de mielina se interrumpe en el nodo de Ranvier. En las fibras amielínicas la propagación del impulso es según la teoría del circuito local; mientras que en las mielínicas la conducción es saltatoria, de un nodo de Ranvier al próximo. Por otra parte las neuronas presentan neurofibrillas en el neuroplasma; estas son el resultado del agrupamiento de neurotúbulos y neurofilamentos. El pericarion es rico en ribosomas y RER (sustancia de Nissl). La abundancia de ribosomas está relacionada con sus funciones biosintéticas. Además de la conducción de impulsos, otra función importante del axón es el transporte axónico de materiales entre el soma y las terminaciones axónicas. En el transporte anterógrado la dirección ocurre desde el cuerpo celular hacia la terminación axónica; en el transporte retrógrado, la dirección es en el sentido contrario. Los microtúbulos son importantes para el transporte anterógrado rápido. El transporte axónico es de importancia crucial para las relaciones tróficas entre las neuronas y los músculos o las glándulas. Si se interrumpen estas relaciones, las células blancos experimentarán atrofia. Se emplea el transporte anterógrado en la translocación de organitos y vesículas, lo mismo que de macromoléculas, como actina, miosina y clatrina, y algunas de las enzimas necesarias para la síntesis de neurotransmisores a nivel de las terminaciones axónicas. Las sustancias que viajan al cuerpo celular por el axón en el transporte retrógrado incluyen proteínas que constituyen la base estructural de los neurofilamentos, subunidades de los microtúbulos y enzimas solubles. También se transportan hacia los endolisosomas del soma moléculas pequeñas y proteínas destinadas a la degradación. Los virus (p. ej., los del herpes simple y de la rabia) pueden valerse del transporte axónico para entrar en una neurona y diseminarse hacia otras entre el cuerpo celular y la terminación nerviosa. En las neuronas se sintetizan numerosos mediadores químicos como la acetilcolina, adrenalina, serotonina, etc. y hormonas, tales como la vasopresina y la oxitocina. La secreción hormonal es característica sólo de neuronas del hipotálamo. En el citoplasma neuronal también tiene lugar la síntesis de glúcidos y lípidos. A diferencia del resto de los tejidos estudiados, en el tejido nervioso el elemento de sostén lo constituyen las denominadas neuroglias, y no los elementos extracelulares fibrosos del tejido conjuntivo. Las neuroglias también desempeñan funciones metabólicas tróficas y de defensa en el tejido nervioso.
August 31, 2008 With Child, With Cancer By PAMELA PAUL LIZETTE IRVIN, HEAVILY PREGNANT , reclined on a hospital bed, relaxed, considering the circumstances. A bag of fluid dripped into her blood through an IV line as Irvin sucked on ice cubes, trying to pass the time. The ice helped to minimize the metallic taste and heat in her mouth from 5- fluorouracil, an antimetabolite, which ent
Aantal in PubMed geciteerde artikelen in 2009 van medische stafleden van het Kennemer Gasthuis, het Spaarne Ziekenhuis en het Streeklaboratorium voor de Volksgezondheid Kennemerland. De vermelding van de artikelen is conform de weergave van de meeste andere STZ-ziekenhuizen: artikelen worden geaccepteerd wanneer de printversie uit is ( ahead of print wordt in het jaardocument 2010 meegenomen);
 TEJIDO NERVIOSO.
TEJIDO NERVIOSO. 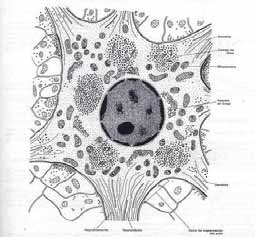
 (transmisor del impulso nervioso), que es uno solo para cada neurona; y las dendritas (receptoras del impulso nervioso), generalmente múltiples. El tamaño del cuerpo o soma de las neuronas varía desde muy pequeño, de 4 a 6 µm, en las llamadas células granulosas o granos del cerebelo, hasta de 150 µm en las células piramidales gigantes de Betz del área motora de la corteza cerebral.
A B Fig. 2. A. Esquema del cuerpo de una neurona multipolar. B, Microfotografía de una neurona multipolar. Cresil violeta. Se observa el nucleolo y los cuerpos de Nissl. La forma de las neuronas también es variada, debido principalmente al número y la disposición de sus prolongaciones. Las neuronas pueden ser estrelladas, fusiformes, piramidales, esféricas, etc. DISTRIBUCIÓN. En el SNC los cuerpos neuronales se agrupan en la corteza cerebral, corteza cerebelosa y en los núcleos grises. Estas zonas en estado fresco presentan un color grisáceo dada la abundancia de cuerpos neuronales y poca presencia de fibras nerviosas mielínicas, a estas zonas se le denomina sustancia gris. En la sustancia gris, además de los somas neuronales y sus prolongaciones, se encuentran gran número de células de neuroglia y capilares sanguíneos. Las zonas del SNC donde predominan las fibras nerviosas mielínicas (axones revestidos de mielina) se les denomina sustancia blanca, ya que por el alto contenido en lípidos de la mielina estas zonas presentan color blanco. En el SNP los cuerpos neuronales se agrupan en los ganglios nerviosos del Sistema Nervioso Autónomo. CLASIFICACIÓN MORFOLÓGICA DE LAS NEURONAS De acuerdo al número de prolongaciones dendriticas las neuronas se clasifican en:
(transmisor del impulso nervioso), que es uno solo para cada neurona; y las dendritas (receptoras del impulso nervioso), generalmente múltiples. El tamaño del cuerpo o soma de las neuronas varía desde muy pequeño, de 4 a 6 µm, en las llamadas células granulosas o granos del cerebelo, hasta de 150 µm en las células piramidales gigantes de Betz del área motora de la corteza cerebral.
A B Fig. 2. A. Esquema del cuerpo de una neurona multipolar. B, Microfotografía de una neurona multipolar. Cresil violeta. Se observa el nucleolo y los cuerpos de Nissl. La forma de las neuronas también es variada, debido principalmente al número y la disposición de sus prolongaciones. Las neuronas pueden ser estrelladas, fusiformes, piramidales, esféricas, etc. DISTRIBUCIÓN. En el SNC los cuerpos neuronales se agrupan en la corteza cerebral, corteza cerebelosa y en los núcleos grises. Estas zonas en estado fresco presentan un color grisáceo dada la abundancia de cuerpos neuronales y poca presencia de fibras nerviosas mielínicas, a estas zonas se le denomina sustancia gris. En la sustancia gris, además de los somas neuronales y sus prolongaciones, se encuentran gran número de células de neuroglia y capilares sanguíneos. Las zonas del SNC donde predominan las fibras nerviosas mielínicas (axones revestidos de mielina) se les denomina sustancia blanca, ya que por el alto contenido en lípidos de la mielina estas zonas presentan color blanco. En el SNP los cuerpos neuronales se agrupan en los ganglios nerviosos del Sistema Nervioso Autónomo. CLASIFICACIÓN MORFOLÓGICA DE LAS NEURONAS De acuerdo al número de prolongaciones dendriticas las neuronas se clasifican en:
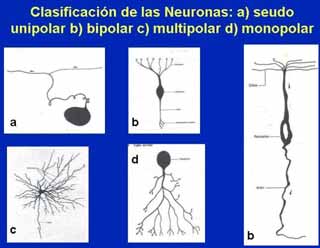
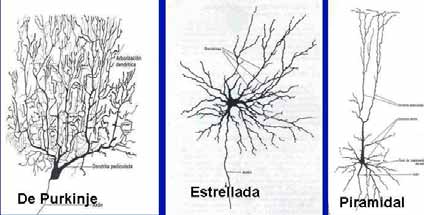 Fig. 3 Clasificación de las neuronas según el número de dendritas.
Fig. 3 Clasificación de las neuronas según el número de dendritas. 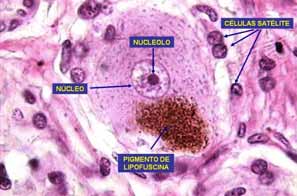
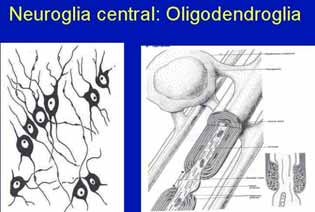 Oligodendroglia. Se parecen a los astrocitos, pero son más pequeños y contienen
Oligodendroglia. Se parecen a los astrocitos, pero son más pequeños y contienen 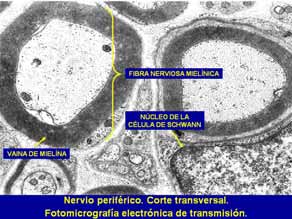
 Al M/O la vaina de mielina aparece como un tubo alongado que se interrumpe a intervalos regulares en los denominados nodos de Ranvier. Los segmentos de mielina entre nudos consecutivos se denominan internodos. En el SNC cada segmento de mielina está formado por una prolongación citoplasmática de una oligodendroglia que se dirige hacia el axón y da varias vueltas alrededor del mismo, enrollándolo con una vaina formada por capas yuxtapuestas de la membrana celular, la vaina de mielina. No está claro el mecanismo de la mielinización, proceso por medio del cual la célula de Schwann (o el oligodendrocito) envuelve su membrana de manera concéntrica alrededor del axón para formar la vaina de mielina. Se cree que se inicia conforme la célula de Schwann va envolviendo al axón y, de alguna manera, envuelve también su membrana alrededor de éste. La envoltura puede proseguir por más de 50 vueltas. Conforme la membrana se enrolla alrededor del axón, produce una serie de líneas densas amplias alternantes con líneas menos densas y más estrechas a intervalos de 12 nm. La microscopía electrónica de alta resolución ha revelado un pequeño espacio dentro de la línea interperiódica entre cada capa espiral de la vaina de mielina, que se denomina espacio interperiódico. Se considera que este espacio ofrece acceso a las pequeñas moléculas para que lleguen al axón. La región de la línea interperiódica que está en contacto íntimo con el axón se conoce como mesoaxón interno, en tanto que su superficie más externa, en contacto con la célula de Schwann es el mesaron externo.
A B Fig. 7. Nervio periférico. A microfotografía electrónica. B microscopía óptica. La microscopía de luz ha revelado diversos surcos oblicuos de forma cónica en la vaina de mielina de cada segmento internodal que se denominan hendiduras de Schmidt-Lantermann. Se demostró que estas hendiduras, vistas con el microscopio electrónico, eran citoplasma de las células de Schwann atrapado dentro de las vainas de mielina. Un axón adquiere su vaina mielínica al final del segmento inicial, es decir, un poco después del cono axónico, y continúa hasta cerca de la terminación axónica. Las porciones desnudas de los nodos de Ranvier son regiones altamente especializadas, responsables de la capacidad autorregenerativa del potencial de acción conducido. Durante el paso de un potencial de acción cambia la conductancia de la membrana entre los nudos, de manera que la onda de despolarización "salta" de nodo a nodo y forma la
Al M/O la vaina de mielina aparece como un tubo alongado que se interrumpe a intervalos regulares en los denominados nodos de Ranvier. Los segmentos de mielina entre nudos consecutivos se denominan internodos. En el SNC cada segmento de mielina está formado por una prolongación citoplasmática de una oligodendroglia que se dirige hacia el axón y da varias vueltas alrededor del mismo, enrollándolo con una vaina formada por capas yuxtapuestas de la membrana celular, la vaina de mielina. No está claro el mecanismo de la mielinización, proceso por medio del cual la célula de Schwann (o el oligodendrocito) envuelve su membrana de manera concéntrica alrededor del axón para formar la vaina de mielina. Se cree que se inicia conforme la célula de Schwann va envolviendo al axón y, de alguna manera, envuelve también su membrana alrededor de éste. La envoltura puede proseguir por más de 50 vueltas. Conforme la membrana se enrolla alrededor del axón, produce una serie de líneas densas amplias alternantes con líneas menos densas y más estrechas a intervalos de 12 nm. La microscopía electrónica de alta resolución ha revelado un pequeño espacio dentro de la línea interperiódica entre cada capa espiral de la vaina de mielina, que se denomina espacio interperiódico. Se considera que este espacio ofrece acceso a las pequeñas moléculas para que lleguen al axón. La región de la línea interperiódica que está en contacto íntimo con el axón se conoce como mesoaxón interno, en tanto que su superficie más externa, en contacto con la célula de Schwann es el mesaron externo.
A B Fig. 7. Nervio periférico. A microfotografía electrónica. B microscopía óptica. La microscopía de luz ha revelado diversos surcos oblicuos de forma cónica en la vaina de mielina de cada segmento internodal que se denominan hendiduras de Schmidt-Lantermann. Se demostró que estas hendiduras, vistas con el microscopio electrónico, eran citoplasma de las células de Schwann atrapado dentro de las vainas de mielina. Un axón adquiere su vaina mielínica al final del segmento inicial, es decir, un poco después del cono axónico, y continúa hasta cerca de la terminación axónica. Las porciones desnudas de los nodos de Ranvier son regiones altamente especializadas, responsables de la capacidad autorregenerativa del potencial de acción conducido. Durante el paso de un potencial de acción cambia la conductancia de la membrana entre los nudos, de manera que la onda de despolarización "salta" de nodo a nodo y forma la
 conducción "saltatoria", que es mucho más rápida que en las fibras amielínicas.
conducción "saltatoria", que es mucho más rápida que en las fibras amielínicas.