He encontrado que alguna farmacia puede tener existencias limitadas de ciertos medicamentos, mientras que otras pueden tener casi cualquier formato que se le ocurra y el habitual de dosis habitualidad apareció. En resumen, siempre se contiene el almacén de corroborar. Al mismo tiempo que el producto que más que gustaba ha resultado no estaba disponible en stock otro distinto por las Buenas costumbres también debe buscarse jefe no asн parezca. Por eso es importante disponer de un Plan B para actuar cuandod ello no ocurra.
Ventaja de tomar un genérico en lugar de Asix
Un genérico es más barato que el nombre de marca
Uno de los mayores incentivos para someterse al Dónde comprar Lasix genérico en lugar de pagar la marca es que usted puede obtener un ahorrando importantes Lasix genérico. Por lo tanto, un Lasix genérico es en general mucho más barato que el homólogo de marca, así que una denominación genérica se hace posible para las personas que usan este medicamento con frecuencia. Un ejemplo: La compra de lurosemida en lugar de Lasix es una considerable ahorro para el presupuesto mensual de medicamentos.
Microsoft word - 2011.3.178-183.doc
AAB BIOFLUX Advances in Agriculture & Botanics- International Journal of the Bioflux Society Effects of cold stress on some Apricot (Prunus armeniaca L.) cultivars in different phenological stages Mehdi Rouhani Nia, Alireza Motallebi-Azar, and Habib Davati-Kazemnia
Department of Horticultural Sciences, Faculty of Agriculture, University of Tabriz,
Corresponding author: A. Motallebi-Azar, [email protected]Abstract. One of the main problems in fruit production is the ambient low temperatures. It means that low temperature in early spring compromised the flowers and fruits. Apricot tree is known as an early blooming and sensitive to frost. In accordance to this in East Azarbaijan which is one of the important apricot production regions in Iran, rate of frost resistance of a few commercial apricot cultivars as Nassiri, Ghermez Shahrood, Dorosht Malaier, Ghorban Maraghe and Kanino were investigated. Experimental samples were studied under four different cold treatment levels (+2, 0, -2, -40C). Experimental traits include sensitivity and difference of anther, petal, stigma, style and ovary to temperature treatment, which have been morphologically evaluated. About all cultivars with progress of phenological stages and temperature decrease, damage percent increased. Among the above cultivars, Dorosh Malaier had a lowest and Ghorban Maraghe had a highest damage percent in low temperature. Nassiri, Ghermez Shahrood and Kanino were palced among mentioned cultivars. Proline quantity after cold treatment was evaluated as well. The results showed that between increase in proline and cold resistance in above mentioned cultivars don’t have a complete liner correlation. Key Words: Apricot, cold stress, phenological stages, proline. Introduction. One of the most important fruit trees in Iran, especially in Azerbaijan region is the Apricot (Prunus armeniaca L.) whose products are used as fresh fruits, jam, can, etc. It's also exported to other countries. Apricot contains an important amount of K+, vitamin A, P, aspartic acid, glutamine and metionine (Tamassy & Zayan 1983). In according to early flowering, cold stress injury is a limiting factor in planting and processing apricots, so in capable areas this factor cause production decreases. Chilling also decrease quality and quantity of products (Probesting 1978). This damage is mostly occurred thorough ice formation as long as cell survival depends on ice location. Ice formation may take place into or out cells (Hare & Cress 1997). A few factors are important in cold hardiness of flower buds, including genotype, flower phonological stages, ice formation place, water content and nutrition (Lu & Rieyer 1993; Rodrigo 2000). In late summer receptors in leaf get signals of shortening the days and produce substances in response which motivated cold hardening. These substances are transferred from leafy branches to leafless ones. This stage is known as primary stage of hardening (Rodrigo 2000). Studies show that leafy branches in sunlight are better acclimated than leafless branches (Yelnosky 1979; Erez et al 1998). The second stage of hardening is motivated by low temperature. Most plants achieve the highest rate of cold resistance after they are exposed freezing temperature. This stage is known as third stage of resistance (Faust 1989; Linden 2002). In some case proline, betaine and glycine can increase the osmotic stress. Proline is the most important because of high osmotic pressure and non toxic at high level (Buban & Touri 1985). Growth regulators treatment sometimes causes higher cold resistance. Abscisic acid (ABA) treatment (1-4 mol) causes cold resistance increased with 2 degrees (-14 to - 160C) (Fuchigamiet et al 1971). In this case ABA causes lipid reduction per oxidation of cell membrane, creates membrane stability and save inters cellular proline (Chen & Li
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
2002). Referring to early flowering of apricot trees in places like East Azarbaijan which are capable of chilling, some part of production is lost annually, so to indicate resistant cultivars five commercial cultivars planted in this area named: Nassiri, Germez Shahrood, Gorbane Maraghe, Kanino and Doroshte Malaier were evaluated. Material and Method. For these study five cultivars from Agricultural Research Center of East Azerbaijan was selected. Three trees were selected from each cultivar. Each experimental units consisting by three branches, each one contains ten buds, i.e. 30 buds for each treatment in four thermal levels (+2, 0, -2, -40C) and six phenological stages (swelled bud, visible petals, visible sepals, full bloom, after petal fall, after fruit set) was studied. After thermal treatment for 16-24 h, samples where placed at room temperature to observe chilling injuries. Firstly anthers, petals, sepals, style, pistil and ovary morphologically were evaluated. In the second stage were evaluated the proline levels by Bietes et al (1973) method. This study was conducted as a multi factorial experiment (5×6×4) in three randomized replication. First factor is the cultivar, second is the phenological stages and third factor is the thermal levels. Data evaluated with MSTATC at 5% level. Results and Discussion. Flowers are sensitive to low temperatures and at long exposure increase the damage percentage. Pistil is more sensitive to stress, i.e. cold stress causes extreme damage in them. Among pistil organs, ovary is more sensitive than style and stigma. Afrigore effects in ovary decreases meristematic activity (Rodrigo 2000).
As it’s shown in Table 1, +20C doesn't damage any flower organs, but decreasing
temperature from +20C to 00C and -20C cause gradually increasing damage. From -20C to -40C the highest damage was observed.
Studies show that increasing of anoxia cause such damages (Probesting 1978).
Some researchers believe that low temperature causes protein breakage which is important for protein shortage and accumulation of toxic products (Hare & Cress 1997).
Studies reports, that survival at low temperature of some cultivars is related to
cell membrane structure and higher amount of unsaturated fatty acids. Some cultivars have supercooling ability which prevents cell damage at lower temperature. Ice formation prevents the trees from supercooling. Other cultivars such as Prunus padus don't have supercooling ability but do to raceme inflorescence some flower survive (Kadir & Probesting 1994).
Apple browning is a distinctive morphological index causing bud drying and their
abscission. Flower outer organs such as petals and sepals may be safe while inner organs such as anthers and ovary could be damaged.
Damaged organs seem brown or yellow-brown which is distinctive morphological
sign of chilling injury (Figure 1). Damage percentage is not only influenced by temperature but is also by phenological stage too. It means that dormant buds have the highest cold resistance, which is related to increasing of inhibitors, decrease of water content and scale formation on the bud at this time. Also in resistant buds a large number of ices is formed in scales and axils, but in non-resistant buds ice formation is inside of flower and bud scales (Rodrigo 2000).
Some cellular changes such as activation of certain genes in low temperature
cause hardening. For example gene Cor-15-a activated in chloroplasts of Arabidopsis thaliana causes freezing preservation of organs (Warren 1998). In alfalfa Ca2+ is transported from cell wall to aqueous part of cytoplasm in cold stress. In addition cold temperature motivates phosphorylation degree changing of some proteins which exist in cytosol. Both of mentioned alteration cause cell survives in cold condition.
Studies show that protein phosphorylation has a role in cold signal transmission.
In such case kinase activity and inactivation of phosphatase cause easier shift of cold signals to nucleus. In alfalfa PP1a and PP2a proteins are responsible for more than 85% cell phosphatase activity (Monory et al 1998). With progress of phenological stages buds are more sensitive, as in full bloom they are more susceptible and the most damage is occurred. The second vulnerable stage is after petal fall. This damage is related to special
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
cell situation and cell wall condition (Table 2). In some species ovary may be safe in cold temperature but anthers or petals may be damaged. In peaches ovary browning may have no external symptoms (Rodrigo 2000). In accordance to high ovary sensitivity it is expected to be the damaged organ while other flower organs to be probably safe (Figure 2).
Damaged flower organs percentage at different temperatures of studied cultivars
*0 indicates no damage, **Means in each column with same letters are not significantly at 5% level, DNMRT.
Damaged flower organs percentage at different phenological stages of studied cultivars
*Samples exposed only at 0 and -20C, ** Means in each column with same letters are not significantly at 5% level, DNMRT. Proline production increases in stress condition. At tall plants are two cycles for proline production: Glotamate and Ornitine. The first cycle is activated by stress or nitrogen
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
shortage and the second one is activated by the high nitrogen level in the cell (Chen & Li 2002).
Proline production preserves NADP+/NADPH levels which have an important role in
electron conduction. In this case increasing oxidative pentose-phosphate cycle activity causes secondary metabolite production (Hare & Cress 1997). In chilling sensitive plants proline increasing is lower than the necessary amount for cold hardiness, unless high level of proline is added before stress (Chen & Li 2002). Of course higher proline level doesn't always cause cold resistance (Yelenosky 1979). Proline production prevents cell from extra acidification.After stress proline catalyses which help mitochondrial oxidative phosphorylation and ATP production helps damage recovery (Hare & Cresss 1997).
In the present study proline samples were evaluated after cold treatment. As
shown in (Table 3) the highest proline level is in winter buds and the lowest in full blooms which is correlated whit damage percent. After petals fall, the proline level is high but newly developed cells have low ability against stress (Table 3). After cold treatment proline increases.
In winter, wheat thermal treatment causes to increase proline. Resistant cultivars
has higher proline rather than sensitive cultivars but there are some exceptions such as 6022w1-121 semi tender winter wheat which has higher proline rather than AF93-2 as a resistant cultivar (Petcu & Terbea 1995).
In studied cultivar Kanino which produces higher proline rather than Dorosht
Malaier, has higher damage percentage too.
Salt treatment in some herbal plants produced higher proline and cold resistance
In resistant Black Locust cultivar, 17 proteins were found that didn't exist in
sensitive cultivars (Tamassy & Zayan 1983).
This study showed that genotype affected damage rate is the lowest at Dorosht
Malayer and the highest at Gorban Maraghe. Other cultivars like Nassiri, Kanino and Germez Shahrood stood between the two motioned cultivars (Table 4).
Yelenosky (1979) conducted that in citrus is no linear correlation in proline level
and cold resistance. Our studies conducted at the same result.
Proline level (micro mol/fw-1) at different phonological stages of studied cultivars
*Means in each column with the same letters are not significantly different at 5% level, DNMRT.
Damage percent of flower organs among studied cultivars
*Means in each column with the same letters are not significantly different at 5% level, DNMRT.
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
Conclusions. Study results showed that cultivars should be selected based on climacteric conditions. In regions with probable spring chilling planting of sensitive cultivars such as Gorban Maraghe should be avoided. On the other hand late irrigation is useful to decrease chilling risk, and irregular fertilization increase chilling damage rate. The best method to increase cold hardiness is to use resistant rootstock such as Harcot, Harvard and Harogen in apricot (Warren 1998). Cultivars with longer dormancy period are recommended. Methods such as mist, fog and sprinkler irrigation have lower effect on chilling preservation.
Not at list growth inhibitors usage leads to improvement of cold hardiness (Mauk
et al 1987). References Bietes S., Waldren R. P., Teare I., 1973 Rapid determination of free proline for water-
stress studies. Plant Soil 39:205-207.
Buban T., Turi I., 1985 Delaying bloom in apricot and peach trees. Acta. Hort. 192:57-
Chen W. P., Li P.H., 2002 Membrane stabilization by abscisic acid under cold aids proline
in alleviating chilling injury in maize (Zea mays L.) cultured cells. Plant Cell Environ 25:955-962.
Erez A., Faust M., Line M., 1998 Chang in water status in peach bud on induction,
development and release from dormancy. Scientia Hortic 73:111-123.
Faust M., 1989 Physiology of temperate zone fruit trees. A Wiley Inter Science Pub. pp:
Fuchigami L. H., Evert R. D., Weiser J. C., 1971 A translocatable cold hardiness
promoter. Plant Physiol 47:164-1.
Hare P. D., Cress W. A., 1997 Metabolic implications of stress-induced proline
accumulation in plants. Plant Growth Regul 21(2):79-102.
Kadir S. A., Probesting E. L., 1994 Various freezing strategies of flower-bud hardiness in
Prunus. J Amer Soc Hort Sci 119(3):584-588.
Linden L., 2002 Measuring cold hardiness in woody plants. PhD Thesis, University of
Helsinky, Department of Applied Biology, 57 pages.
Lu S., Rieyer M., 1993 Effect of temperature preconditioning on ovary frizzing tolerance
of fully opened peach flower. J Hort Sci 68:343-347.
Mauk C. S., Bausher M. G., Yelenosky G., 1987 Physiological effects of temperature and
growth regulators on foliar chlorophyll, soluble proteins and cold hardiness in citrus. Plant Growth Regulation 5(2)141-154.
Monory F., Sanywan V., Dhindsa S., 1998 Low temperature signal transduction during
cold acclimation: Protein phosphates 2A as an early target for cold inactivation. Plant J 13:653-660.
Petcu E., Terbea M., 1995 Proline content and the conductivity test as screening methods
for frost tolerance of winter wheat. Plant Physiol 21:3-11.
Probesting E. L., 1978 Adapting cold hardiness concepts to deciduous fruit culture. In:
Plant cold hardiness and freezing stress. Li P.H., Sakai A. (ed), pp. 267-281, Academic Press Inc.
Rodrigo J., 2000 Spring frost in deciduous fruit trees morphological damage and flower
hardiness. Scientia Hortic 85:155-173.
Ryu B., Costa A., Li P. H., 1995 Induction of cold hardiness by salt-stress involves
synthesis of cold and abscisic acid responsive proteins in potato. Plant Cell Physiol 36:1245-1251.
Tamassy I., Zayan M., 1983 Soluble proteins and amino acids as related to cold
hardiness in some apricot cultivars belonging to different groups. Acta Hort 121:141-153.
Warren J. G., 1998 Manipulating freezing tolerance in plants. Current Biology 8:514-516. Yelnosky G., 1979 Accumulation of free proline in citrus leaves during cold hardening of
young tree in controlled temperature regimes. Plant Physiol 64:425-427.
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
Received: 17 December 2011. Accepted: 18 December 2011. Published online: 23 December 2011. Authors: Mehdi Rouhani Nia, Department of Horticultural Sciences, Faculty of Agriculture, University of Tabriz, 5166616471, Tabriz, Iran, e-mail: [email protected] Alireza Motallebi-Azar, Department of Horticultural Sciences, Faculty of Agriculture, University of Tabriz, 5166616471, Tabriz, Iran, e-mail: [email protected] Habib Davati-Kazemnia, Department of Horticultural Sciences, Faculty of Agriculture, University of Tabriz, 5166616471, Tabriz, Iran, e-mail: [email protected] How to cite this article: Nia M. R., Motallebi-Azar A., Davati-Kazemnia H., 2011 Effects of cold stress on some Apricot (Prunus armeniaca L.) cultivars in different phenological stages. AAB Bioflux 3(3):178-183.
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
Cuadernos del CieJ Cuadernos del CieJ Serie Conferencias Cuadernos del CieJ Detrás de cada "Cuaderno del CIEJ" se encuentra el esfuerzo de mucha gente. Desde los autores que le han dedicado su tiempo desinteresada mente, hasta los compañeros que hacen posible la publicación. Los que desgraban las cintas y tipean las ponencias, los que cuidan la ed
Monday, 16th of November 2009 Workshop - Photogrammetry - Documentation from Photos Workshop - Digital documentation in archaeology Workshop - "Newbies"N. HIRTL / P. DORNINGER, Austria Terrestrial Laser Scanning for accurate restoration documentationKatharina HOLZINGER / Christian SAFRAN / Martin EBNER / Andreas HOLZINGER, Austria Geo-Tagging in Archaeology: Practical Experiences wit

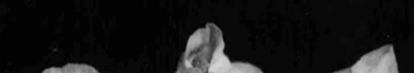
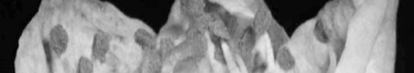



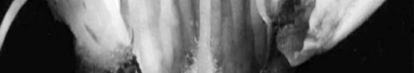
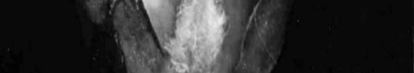











 cell situation and cell wall condition (Table 2). In some species ovary may be safe in cold temperature but anthers or petals may be damaged. In peaches ovary browning may have no external symptoms (Rodrigo 2000). In accordance to high ovary sensitivity it is expected to be the damaged organ while other flower organs to be probably safe (Figure 2).
Damaged flower organs percentage at different temperatures of studied cultivars
*0 indicates no damage, **Means in each column with same letters are not significantly at 5% level, DNMRT.
Damaged flower organs percentage at different phenological stages of studied cultivars
*Samples exposed only at 0 and -20C, ** Means in each column with same letters are not significantly at 5% level, DNMRT. Proline production increases in stress condition. At tall plants are two cycles for proline production: Glotamate and Ornitine. The first cycle is activated by stress or nitrogen
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
shortage and the second one is activated by the high nitrogen level in the cell (Chen & Li 2002).
Proline production preserves NADP+/NADPH levels which have an important role in
electron conduction. In this case increasing oxidative pentose-phosphate cycle activity causes secondary metabolite production (Hare & Cress 1997). In chilling sensitive plants proline increasing is lower than the necessary amount for cold hardiness, unless high level of proline is added before stress (Chen & Li 2002). Of course higher proline level doesn't always cause cold resistance (Yelenosky 1979). Proline production prevents cell from extra acidification.After stress proline catalyses which help mitochondrial oxidative phosphorylation and ATP production helps damage recovery (Hare & Cresss 1997).
In the present study proline samples were evaluated after cold treatment. As
shown in (Table 3) the highest proline level is in winter buds and the lowest in full blooms which is correlated whit damage percent. After petals fall, the proline level is high but newly developed cells have low ability against stress (Table 3). After cold treatment proline increases.
In winter, wheat thermal treatment causes to increase proline. Resistant cultivars
has higher proline rather than sensitive cultivars but there are some exceptions such as 6022w1-121 semi tender winter wheat which has higher proline rather than AF93-2 as a resistant cultivar (Petcu & Terbea 1995).
In studied cultivar Kanino which produces higher proline rather than Dorosht
Malaier, has higher damage percentage too.
Salt treatment in some herbal plants produced higher proline and cold resistance
In resistant Black Locust cultivar, 17 proteins were found that didn't exist in
sensitive cultivars (Tamassy & Zayan 1983).
This study showed that genotype affected damage rate is the lowest at Dorosht
Malayer and the highest at Gorban Maraghe. Other cultivars like Nassiri, Kanino and Germez Shahrood stood between the two motioned cultivars (Table 4).
Yelenosky (1979) conducted that in citrus is no linear correlation in proline level
and cold resistance. Our studies conducted at the same result.
Proline level (micro mol/fw-1) at different phonological stages of studied cultivars
*Means in each column with the same letters are not significantly different at 5% level, DNMRT.
Damage percent of flower organs among studied cultivars
*Means in each column with the same letters are not significantly different at 5% level, DNMRT.
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
Conclusions. Study results showed that cultivars should be selected based on
cell situation and cell wall condition (Table 2). In some species ovary may be safe in cold temperature but anthers or petals may be damaged. In peaches ovary browning may have no external symptoms (Rodrigo 2000). In accordance to high ovary sensitivity it is expected to be the damaged organ while other flower organs to be probably safe (Figure 2).
Damaged flower organs percentage at different temperatures of studied cultivars
*0 indicates no damage, **Means in each column with same letters are not significantly at 5% level, DNMRT.
Damaged flower organs percentage at different phenological stages of studied cultivars
*Samples exposed only at 0 and -20C, ** Means in each column with same letters are not significantly at 5% level, DNMRT. Proline production increases in stress condition. At tall plants are two cycles for proline production: Glotamate and Ornitine. The first cycle is activated by stress or nitrogen
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
shortage and the second one is activated by the high nitrogen level in the cell (Chen & Li 2002).
Proline production preserves NADP+/NADPH levels which have an important role in
electron conduction. In this case increasing oxidative pentose-phosphate cycle activity causes secondary metabolite production (Hare & Cress 1997). In chilling sensitive plants proline increasing is lower than the necessary amount for cold hardiness, unless high level of proline is added before stress (Chen & Li 2002). Of course higher proline level doesn't always cause cold resistance (Yelenosky 1979). Proline production prevents cell from extra acidification.After stress proline catalyses which help mitochondrial oxidative phosphorylation and ATP production helps damage recovery (Hare & Cresss 1997).
In the present study proline samples were evaluated after cold treatment. As
shown in (Table 3) the highest proline level is in winter buds and the lowest in full blooms which is correlated whit damage percent. After petals fall, the proline level is high but newly developed cells have low ability against stress (Table 3). After cold treatment proline increases.
In winter, wheat thermal treatment causes to increase proline. Resistant cultivars
has higher proline rather than sensitive cultivars but there are some exceptions such as 6022w1-121 semi tender winter wheat which has higher proline rather than AF93-2 as a resistant cultivar (Petcu & Terbea 1995).
In studied cultivar Kanino which produces higher proline rather than Dorosht
Malaier, has higher damage percentage too.
Salt treatment in some herbal plants produced higher proline and cold resistance
In resistant Black Locust cultivar, 17 proteins were found that didn't exist in
sensitive cultivars (Tamassy & Zayan 1983).
This study showed that genotype affected damage rate is the lowest at Dorosht
Malayer and the highest at Gorban Maraghe. Other cultivars like Nassiri, Kanino and Germez Shahrood stood between the two motioned cultivars (Table 4).
Yelenosky (1979) conducted that in citrus is no linear correlation in proline level
and cold resistance. Our studies conducted at the same result.
Proline level (micro mol/fw-1) at different phonological stages of studied cultivars
*Means in each column with the same letters are not significantly different at 5% level, DNMRT.
Damage percent of flower organs among studied cultivars
*Means in each column with the same letters are not significantly different at 5% level, DNMRT.
AAB Bioflux, 2011, Volume 3, Issue 3. http://www.aab.bioflux.com.ro
Conclusions. Study results showed that cultivars should be selected based on